Embryonale Homologien
Eine der wichtigsten Unterscheidungen der evolutionären Embryologen war der Unterschied zwischen Analogie und Homologie. Beide Begriffe beziehen sich auf Strukturen, die ähnlich zu sein scheinen. Homologe Strukturen sind jene Organe, deren zugrunde liegende Ähnlichkeit sich daraus ergibt, dass sie von einer gemeinsamen Ahnenstruktur abgeleitet sind. Zum Beispiel sind der Flügel eines Vogels und das Vorderglied eines Menschen homolog. Darüber hinaus sind ihre jeweiligen Teile homolog (Abbildung 1.13)., Analoge Strukturen sind solche, deren Ähnlichkeit von ihrer Ausführung einer ähnlichen Funktion herrührt, anstatt von einem gemeinsamen Vorfahren. Daher sind beispielsweise der Flügel eines Schmetterlings und der Flügel eines Vogels analog. Die beiden Flügeltypen haben eine gemeinsame Funktion (und werden daher beide Flügel genannt), aber der Vogelflügel und der Insektenflügel entsprachen nicht einer ursprünglichen Ahnenstruktur, die durch die Evolution zu Vogelflügeln und Schmetterlingsflügeln modifiziert wurde.
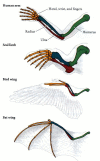
Abbildung 1.,13
Homologien der Struktur unter einem menschlichen Arm, einem Robbenvorarm, einem Vogelflügel und einem Fledermausflügel; homologe Stützstrukturen sind in der gleichen Farbe dargestellt. Alle vier sind homolog als Forelimbs und wurden von einem gemeinsamen Tetrapodenvorfahren abgeleitet. Die Anpassungen (mehr…)
Homologien muss aufmerksam gemacht werden und muss beziehen sich immer auf der Ebene der Organisation verglichen wird. Zum Beispiel sind der Vogelflügel und der Fledermausflügel homolog als Vorderbeine, aber nicht als Flügel., Mit anderen Worten, Sie teilen eine gemeinsame zugrunde liegende Struktur von Vorderbeinknochen, da Vögel und Säugetiere eine gemeinsame Abstammung haben. Der Vogelflügel entwickelte sich jedoch unabhängig vom Fledermausflügel. Fledermäuse stammten von einer langen Reihe von Säugetieren ab, und die Struktur des Fledermausflügels unterscheidet sich deutlich von der eines Vogelflügels.
Einer der berühmtesten Fälle der embryonalen Homologie ist der des Fischkiemenknorpels, des Reptilienkiefers und des Mittelohrs von Säugetieren (rezensiert in Gould 1990)., Zuerst wurden die Kiemenbögen von kieferlosen (Agnathan) Fischen modifiziert, um den Kiefer der Kieferfische zu bilden. In den kieferlosen Fischen öffnete sich eine Reihe von Kiemen hinter dem kieferlosen Mund. Als die Kiemenschlitze durch knorpelige Elemente gestützt wurden, umgab der erste Satz dieser Kiemenstützen den Mund, um den Kiefer zu bilden. Es gibt genügend Beweise dafür, dass Kiefer modifizierte Kiemenstützen sind. Erstens werden diese beiden Knochensätze aus Neuralkammzellen hergestellt. (Die meisten anderen Knochen stammen aus mesodermalem Gewebe.) Zweitens bilden sich beide Strukturen aus oberen und unteren Stäben, die sich nach vorne biegen und in der Mitte aufklappbar sind., Drittens scheint die Kiefermuskulatur homolog zur ursprünglichen Kiemenunterstützungsmuskulatur zu sein. Somit scheint der Wirbeltierkiefer homolog zu den Kiemenbögen von kieferlosen Fischen zu sein.
Aber die Geschichte endet hier nicht. Der obere Teil des zweiten Embryonalbogens, der die Kiemen stützte, wurde zum hyomandibulären Knochen von Kieferfischen. Dieses Element stützt den Schädel und verbindet den Kiefer mit dem Schädel (Abbildung 1.14 A). Als Wirbeltiere an Land kamen, hatten sie ein neues Problem: Wie man in einem Medium hört, das so dünn wie Luft ist., Der hyomandibuläre Knochen befindet sich zufällig in der Nähe der Ohrkapsel, und knöchernes Material eignet sich hervorragend zur Schallübertragung. So funktionierte der hyomandibuläre Knochen der ersten Amphibien, während er noch als Schädelstütze fungierte, auch als Schallwandler (Clack 1989). Als die terrestrischen Wirbeltiere ihre Fortbewegung, Kieferstruktur und Haltung veränderten, wurde der Schädel fest am Rest des Schädels befestigt und brauchte die hyomandibuläre Klammer nicht. Der hyomandibuläre Knochen scheint sich dann auf den Stapes-Knochen des Mittelohrs spezialisiert zu haben., Was die sekundäre Funktion dieses Knochens war, wurde seine primäre Funktion.
Die ursprünglichen Kieferknochen haben sich ebenfalls geändert. Der erste embryonale Bogen erzeugt den Kieferapparat. Bei Amphibien, Reptilien und Vögeln bildet der hintere Teil dieses Knorpels den quadratischen Knochen des Oberkiefers und den Gelenkknochen des Unterkiefers. Diese Knochen verbinden sich miteinander und sind für die Artikulation des Ober-und Unterkiefers verantwortlich. Bei Säugetieren tritt diese Artikulation jedoch an einer anderen Region auf (den D-und Plattenepithelknochen), wodurch diese knöchernen Elemente „befreit“ werden, um neue Funktionen zu erhalten., Der viereckige Knochen des Reptilienoberkiefers entwickelte sich zum Säugetier-Incus-Knochen des Mittelohrs, und der Gelenkknochen des Unterkiefers des Reptils wurde zu unserem Malleus. Dieser letztere Prozess wurde erstmals 1837 von Reichert beschrieben, als er im Schweinembryo beobachtete, dass der Unterkiefer (Kieferknochen) an der Seite von Meckels Knorpel verknöchert, während der hintere Bereich von Meckels Knorpel verknöchert,sich vom Rest des Knorpels löst und in den Bereich des Mittelohrs eintritt, um der Malleus zu werden (Abbildung 1.14 B, C)., So sind die Mittelohrknochen des Säugetiers homolog zum hinteren Unterkiefer des Reptils und zu den Kiemenbögen der Agnathan-Fische. In Kapitel 22 werden neuere Informationen über das Verhältnis von Entwicklung und Evolution beschrieben.