Mikrotubuli
Diese geraden, hohlen Zylinder befinden sich im gesamten Zytoplasma aller eukaryotischen Zellen (Prokaryoten haben sie nicht) und erfüllen eine Vielzahl von Funktionen, die vom Transport bis zur strukturellen Unterstützung reichen. Mikrotubuli mit einem Durchmesser von etwa 25 Nanometern sind Teil des Zytoskeletts, das einer Zelle Struktur und Form verleiht, und dienen auch als Förderbänder, die andere Organellen im gesamten Zytoplasma bewegen., Darüber hinaus sind Mikrotubuli die Hauptbestandteile von Zilien und Flagellen und beteiligen sich an der Bildung von Spindelfasern während der Zellteilung (Mitose). Die Länge der Mikrotubuli in der Zelle variiert zwischen 200 Nanometern und 25 Mikrometern, abhängig von der Aufgabe eines bestimmten Mikrotubuli und dem Zustand des Lebenszyklus der Zelle.
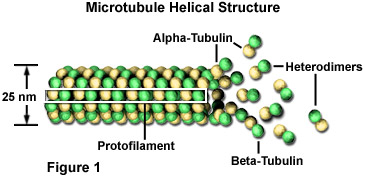
Mikrotubuli sind Biopolymere, die aus Untereinheiten bestehen, die aus einem reichlich vorhandenen kugelförmigen zytoplasmatischen Protein bestehen, das als Tubulin bekannt ist, wie in Abbildung 1 dargestellt., Jede Untereinheit des Mikrotubulus besteht aus zwei leicht unterschiedlichen, aber eng verwandten einfacheren Einheiten, Alpha-Tubulin und Beta-Tubulin, die sehr eng miteinander verbunden sind, um Heterodimer zu bilden. In einem Mikrotubulus sind die Untereinheiten so organisiert, dass sie alle die gleiche Richtung zeigen, um 13 parallele Protofilamente zu bilden. Diese Organisation verleiht der Struktur Polarität, wobei nur die Alpha-Tubulin-Proteine an einem Ende und nur Beta-Tubulin-Proteine am anderen Ende exponiert sind.,
Durch Zugabe oder Entfernung von kugelförmigen Tubulinproteinen kann die Länge polymerer Mikrotubuli erhöht oder verringert werden. Da die beiden Enden eines Mikrotubulus jedoch nicht gleich sind, ist die Geschwindigkeit, mit der Wachstum oder Depolymerisation an jedem Pol auftritt, unterschiedlich. Das Ende eines polarisierten Filaments, das am schnellsten wächst und schrumpft, wird als Plusende und das gegenüberliegende Ende als Minusende bezeichnet. Für alle Mikrotubuli ist das Minus-Ende das mit exponierten Alpha-Tubulinen., In einer tierischen Zelle befindet sich dieses Ende am zentriolhaltigen Zentrosom in der Nähe des Kerns, während das Plus-Ende, das aus exponierten Beta-Einheiten besteht, auf die Oberfläche der Zelle projiziert wird. Mikrotubuli werden kontinuierlich zusammengebaut und demontiert, so dass Tubulinmonomere an anderer Stelle transportiert werden können, um bei Bedarf Mikrotubuli zu bauen.,
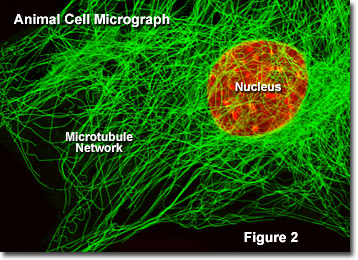
In Abbildung 2 ist ein digitales Bild des Mikrotubuli-Netzwerks, das in einer embryonalen Mauszelle gefunden wird, wie es durch ein optisches Fluoreszenzmikroskop gesehen wird. Das ausgedehnte verflochtene Netzwerk wird mit primären Antikörpern gegen Alpha-Tubulin markiert, die dann mit sekundären Antikörpern gefärbt werden, die einen grün fluoreszierenden Farbstoff enthalten. Der Kern wurde mit einem roten Farbstoff gegengefärbt, um seine Position in Bezug auf das Mikrotubuli-Netzwerk zu notieren., Die Fluoreszenzmikroskopie ist ein wichtiges Instrument, mit dem Wissenschaftler die Struktur und Funktion innerer Zellorganellen untersuchen.
Neben ihrer strukturellen Stützrolle dienen Mikrotubuli auch als Autobahnsystem, auf dem Organellen mit Hilfe motorischer Proteine transportiert werden können. Zum Beispiel verbindet das Mikrotubuli-Netzwerk den Golgi-Apparat mit der Plasmamembran, um sekretorische Vesikel für den Export zu leiten, und transportiert auch Mitochondrien im Zytoplasma hin und her., Ein anderes Beispiel ist die Translokation von Vesikeln, die Neurotransmitter enthalten, durch Mikrotubuli zu den Spitzen von Nervenzellaxonen. Die am Organellentransport beteiligten motorischen Proteine verändern ihre dreidimensionale Konformation unter Verwendung von Adenosintriphosphat (ATP) als Brennstoff, um sich entlang eines Mikrotubulus hin und her zu bewegen. Mit jedem Schritt gibt das Motormolekül einen Teil des Mikrotubulus frei und greift eine zweite Stelle weiter vom Filament. Motorproteine, die in mehrere verschiedene Klassen eingeteilt sind, haften über spezialisierte Rezeptoren an Organellen.,
Da eukaryotische Zellen stark von der Integrität von Mikrotubuli und anderen zytoskeletalen Filamenten abhängen, um ihre Struktur aufrechtzuerhalten und im Wesentlichen zu überleben, produzieren viele Pflanzen natürliche Toxine, die darauf abzielen, das Mikrotubuli-Netzwerk als Mittel zur Selbstverteidigung zu stören. Taxol ist beispielsweise eine toxische Substanz, die von einer Eibenart produziert wird und die die Polymerisation von Mikrotubuli (Aufbau eines Makromoleküls) erhöht, indem sie an das Filament bindet und stabilisiert., Andere natürliche Toxine, wie das vom Wiesensafran produzierte Colchicin, destabilisieren Mikrotubuli und behindern deren Polymerisation. Beide Arten von Ereignissen können für die betroffene Zelle tödlich sein, obwohl dies unter bestimmten Umständen für Tiere von Vorteil sein kann, wie Taxol zeigt, das üblicherweise als Krebsmedikament verwendet wird.
ZURÜCK ZUR TIERISCHEN ZELLSTRUKTUR
ZURÜCK ZUR PFLANZLICHEN ZELLSTRUKTUR
Fragen oder Kommentare? Senden Sie uns eine E-Mail.
© 1995-2019 von Michael W. Davidson und der Florida State University. Alle Rechte Vorbehalten., Keine Bilder, Grafiken, Software, Skripte oder Applets dürfen ohne Erlaubnis der Urheberrechtsinhaber reproduziert oder in irgendeiner Weise verwendet werden. Durch die Nutzung dieser Website stimmen Sie allen von den Eigentümern festgelegten rechtlichen Bedingungen zu.
Diese Website wird von unserem
Graphics & Web Programming Team
in Zusammenarbeit mit Optical Microscopy am
National High Magnetic Field Laboratory gepflegt.,
Last modification: Friday, Nov 13, 2015 at 01:18 PM
Access Count Since October 1, 2000: 282919
Microscopes provided by:
![]()
![]()