Obiettivi di Apprendimento
- Descrivere litico e lisogeni cicli di vita
- Descrivere il processo di replicazione del virus animali
- Descrivere le caratteristiche uniche di retrovirus e virus latenti
- Discutere umano virus e virus-cellula ospite interazioni
- Spiegare il processo di trasduzione
- Descrivere il processo di replicazione del virus della pianta
Tutti i virus dipendono dalle cellule per la riproduzione e processi metabolici., Da soli, i virus non codificano per tutti gli enzimi necessari per la replicazione virale. Ma all’interno di una cellula ospite, un virus può requisire macchinari cellulari per produrre più particelle virali. I batteriofagi si replicano solo nel citoplasma, poiché le cellule procariote non hanno un nucleo o organelli. Nelle cellule eucariotiche, la maggior parte dei virus a DNA può replicarsi all’interno del nucleo, con un’eccezione osservata nei grandi virus a DNA, come i poxvirus, che possono replicarsi nel citoplasma. I virus a RNA che infettano le cellule animali spesso si replicano nel citoplasma.,
Il ciclo di vita dei virus con ospiti procarioti
Il ciclo di vita dei batteriofagi è stato un buon modello per capire come i virus influenzano le cellule che infettano, poiché sono stati osservati processi simili per i virus eucariotici, che possono causare la morte immediata della cellula o stabilire un’infezione latente o cronica. I fagi virulenti in genere portano alla morte della cellula attraverso la lisi cellulare., I fagi temperati, d’altra parte, possono diventare parte di un cromosoma ospite e vengono replicati con il genoma cellulare fino a quando non vengono indotti a creare virus appena assemblati o virus progenie.
Il ciclo litico
Durante il ciclo litico del fago virulento, il batteriofago assume la cellula, riproduce nuovi fagi e distrugge la cellula. Il fago T-even è un buon esempio di una classe ben caratterizzata di fagi virulenti. Ci sono cinque fasi nel ciclo litico batteriofago (vedi Figura 1)., L’attaccamento è la prima fase del processo di infezione in cui il fago interagisce con specifici recettori superficiali batterici (ad esempio, lipopolisaccaridi e proteine Omc sulle superfici ospiti). La maggior parte dei fagi ha una gamma ristretta di ospiti e può infettare una specie di batteri o un ceppo all’interno di una specie. Questo riconoscimento unico può essere sfruttato per il trattamento mirato dell’infezione batterica dalla terapia del fago o per la tipizzazione del fago per identificare le sottospecie o gli sforzi batterici unici. Il secondo stadio dell’infezione è l’ingresso o la penetrazione., Ciò avviene attraverso la contrazione della guaina della coda, che agisce come un ago ipodermico per iniettare il genoma virale attraverso la parete cellulare e la membrana. La testa del fago e i componenti rimanenti rimangono al di fuori dei batteri.

Figura 1. Un fago virulento mostra solo il ciclo litico qui raffigurato. Nel ciclo litico, il fago replica e lisizza la cellula ospite.
Il terzo stadio dell’infezione è la biosintesi di nuovi componenti virali., Dopo essere entrato nella cellula ospite, il virus sintetizza le endonucleasi codificate dal virus per degradare il cromosoma batterico. Quindi dirotta la cellula ospite per replicare, trascrivere e tradurre i componenti virali necessari (capsomeri, guaina, piastre di base, fibre della coda ed enzimi virali) per l’assemblaggio di nuovi virus. I geni della polimerasi sono solitamente espressi all’inizio del ciclo, mentre le proteine del capside e della coda sono espresse in seguito. Durante la fase di maturazione, vengono creati nuovi virioni. Per liberare i fagi liberi, la parete cellulare batterica viene interrotta dalle proteine dei fagi come holin o lisozima., La fase finale è il rilascio. I virus maturi scoppiano dalla cellula ospite in un processo chiamato lisi ed i virus della progenie sono liberati nell’ambiente per infettare le nuove cellule.
Il ciclo lisogenico
In un ciclo lisogenico, il genoma del fago entra anche nella cellula attraverso l’attaccamento e la penetrazione. Un primo esempio di un fago con questo tipo di ciclo di vita è il fago lambda. Durante il ciclo lisogenico, invece di uccidere l’ospite, il genoma del fago si integra nel cromosoma batterico e diventa parte dell’ospite. Il genoma fago integrato è chiamato un profago., Un ospite batterico con un profago è chiamato lisogeno. Il processo in cui un batterio è infettato da un fago temperato è chiamato lisogenesi. È tipico dei fagi temperati essere latenti o inattivi all’interno della cellula. Mentre il batterio replica il suo cromosoma, replica anche il DNA del fago e lo trasmette a nuove cellule figlie durante la riproduzione. La presenza del fago può alterare il fenotipo del batterio, in quanto può portare in geni extra (ad esempio, geni tossina che possono aumentare la virulenza batterica)., Questo cambiamento nel fenotipo ospite è chiamato conversione lisogenica o conversione dei fagi. Alcuni batteri, come Vibrio cholerae e Clostridium botulinum, sono meno virulenti in assenza del profago. I fagi che infettano questi batteri portano i geni della tossina nel loro genoma e migliorano la virulenza dell’ospite quando i geni della tossina sono espressi. Nel caso di V. colera, la tossina codificata fago può causare grave diarrea; in C. botulinum, la tossina può causare paralisi., Durante la lisogenesi, la prophage persisterà nel cromosoma ospite fino all’induzione, che si traduce nell’escissione del genoma virale dal cromosoma ospite. Dopo che si è verificata l’induzione, il fago temperato può procedere attraverso un ciclo litico e quindi sottoporsi a lisogenesi in una cellula appena infetta (vedere Figura 2).

Figura 2. Un batteriofago temperato ha cicli litici e lisogenici. Nel ciclo lisogenico, il DNA dei fagi è incorporato nel genoma ospite, formando un profago, che viene trasmesso alle generazioni successive di cellule., Fattori di stress ambientali come la fame o l’esposizione a sostanze chimiche tossiche possono causare l’asportazione del profago ed entrare nel ciclo litico.
Questo video illustra le fasi del ciclo di vita lisogenico di un batteriofago e il passaggio ad una fase litica.
Pensaci
- Un fago latente non è rilevabile in un batterio?
Trasduzione
La trasduzione si verifica quando un batteriofago trasferisce il DNA batterico da un batterio all’altro durante infezioni sequenziali., Esistono due tipi di trasduzione: trasduzione generalizzata e specializzata. Durante il ciclo litico della replicazione virale, il virus dirotta la cellula ospite, degrada il cromosoma ospite e produce più genomi virali. Mentre assembla e confeziona il DNA nella testa del fago, l’imballaggio occasionalmente commette un errore. Invece di impacchettare il DNA virale, prende un pezzo casuale di DNA ospite e lo inserisce nel capside. Una volta rilasciato, questo virione inietterà il DNA dell’ospite precedente in un ospite appena infetto., Il trasferimento asessuato di informazioni genetiche può consentire la ricombinazione del DNA, fornendo così al nuovo ospite nuovi geni (ad esempio, un gene di resistenza agli antibiotici o un gene che metabolizza lo zucchero).
La trasduzione generalizzata si verifica quando un pezzo casuale di DNA cromosomico batterico viene trasferito dal fago durante il ciclo litico. La trasduzione specializzata si verifica alla fine del ciclo lisogenico, quando il profago viene asportato e il batteriofago entra nel ciclo litico. Poiché il fago è integrato nel genoma ospite, il prophage può replicarsi come parte dell’ospite., Tuttavia, alcune condizioni (ad esempio, esposizione alla luce ultravioletta o esposizione chimica) stimolano il prophage a subire induzione, causando il fago di asportare dal genoma, entrare nel ciclo litico, e produrre nuovi fagi di lasciare le cellule ospiti. Durante il processo di escissione dal cromosoma ospite, un fago può occasionalmente rimuovere del DNA batterico vicino al sito di integrazione virale. Il DNA del fago e dell’ospite da un’estremità o da entrambe le estremità del sito di integrazione sono impacchettati all’interno del capside e vengono trasferiti al nuovo ospite infetto., Poiché il DNA trasferito dal fago non è confezionato in modo casuale ma è invece un pezzo specifico di DNA vicino al sito di integrazione, questo meccanismo di trasferimento genico è indicato come trasduzione specializzata (vedi Figura 3). Il DNA può quindi ricombinarsi con il cromosoma ospite, dando a quest’ultimo nuove caratteristiche. La trasduzione sembra svolgere un ruolo importante nel processo evolutivo dei batteri, dando loro un meccanismo per lo scambio asessuato di informazioni genetiche.
Figura 3., Questo diagramma di flusso illustra il meccanismo della trasduzione specializzata. Un fago integrato eccede, portando con sé un pezzo di DNA adiacente al suo punto di inserimento. Sulla reinfezione di un nuovo batterio, il DNA dei fagi si integra con il materiale genetico acquisito dall’ospite precedente.
Pensaci
- Quale ciclo di vita dei fagi è associato a quali forme di trasduzione?,
Ciclo di vita dei virus con ospiti animali
I virus animali litici seguono fasi di infezione simili ai batteriofagi: attacco, penetrazione, biosintesi, maturazione e rilascio (vedi Figura 4). Tuttavia, i meccanismi di penetrazione, biosintesi dell’acido nucleico e rilascio differiscono tra virus batterici e animali. Dopo il legame con i recettori ospiti, i virus animali entrano attraverso l’endocitosi (inghiottimento dalla cellula ospite) o attraverso la fusione della membrana (involucro virale con la membrana cellulare ospite)., Molti virus sono specifici dell’ospite, il che significa che infettano solo un certo tipo di ospite; e la maggior parte dei virus infetta solo alcuni tipi di cellule all’interno dei tessuti. Questa specificità è chiamata tropismo tissutale. Esempi di ciò sono dimostrati dal poliovirus, che presenta tropismo per i tessuti del cervello e del midollo spinale, o dal virus dell’influenza, che ha un tropismo primario per le vie respiratorie.

Figura 4. Nell’infezione da virus influenzale, le glicoproteine virali attaccano il virus a una cellula epiteliale ospite. Di conseguenza, il virus è inghiottito., RNA virale e proteine virali sono fatti e assemblati in nuovi virioni che vengono rilasciati dal germogliamento.
I virus animali non sempre esprimono i loro geni usando il normale flusso di informazioni genetiche-dal DNA all’RNA alle proteine. Alcuni virus hanno un genoma dsDNA come organismi cellulari e possono seguire il flusso normale. Tuttavia, altri possono avere genomi ssDNA, dsRNA o ssRNA. La natura del genoma determina come il genoma viene replicato ed espresso come proteine virali., Se un genoma è ssDNA, gli enzimi ospiti saranno utilizzati per sintetizzare un secondo filamento che è complementare al filamento del genoma, producendo così dsDNA. Il dsDNA può ora essere replicato, trascritto e tradotto in modo simile al DNA ospite.
Se il genoma virale è RNA, deve essere utilizzato un meccanismo diverso. Esistono tre tipi di genoma dell’RNA: dsRNA, RNA a singolo filamento positivo ( + ) (+ssRNA) o negativo (-) a singolo filamento (−ssRNA). Se un virus ha un genoma + ssRNA, può essere tradotto direttamente per produrre proteine virali. Genomico virale + ssRNA agisce come mRNA cellulare., Tuttavia, se un virus contiene un genoma a-ssRNA, i ribosomi ospiti non possono tradurlo fino a quando il-ssRNA non viene replicato in + ssRNA dalla RNA polimerasi RNA-dipendente virale (RdRP) (vedere Figura 5). Il RdRP è portato dal virus e può essere usato per fare +ssRNA dal genoma originale −ssRNA. L’RdRP è anche un enzima importante per la replicazione dei virus dsRNA, perché utilizza il filamento negativo del genoma a doppio filamento come modello per creare +ssRNA. Le copie di nuova sintesi +ssRNA possono quindi essere tradotte dai ribosomi cellulari.,
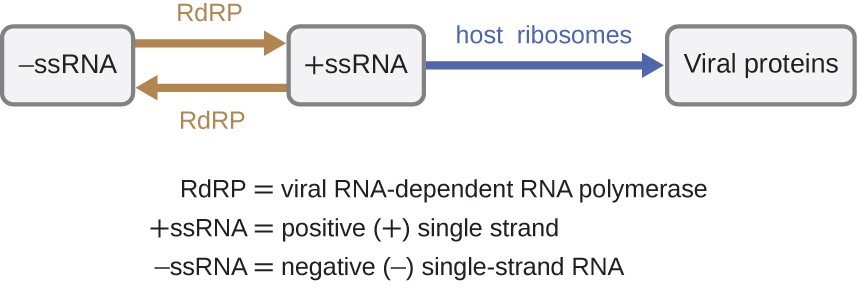
Figura 5. I virus a RNA possono contenere + ssRNA che possono essere letti direttamente dai ribosomi per sintetizzare le proteine virali. I virus contenenti-ssRNA devono prima utilizzare il-ssRNA come modello per la sintesi di +ssRNA prima che le proteine virali possano essere sintetizzate.
Un meccanismo alternativo per la sintesi dell’acido nucleico virale è osservato nei retrovirus, che sono virus +ssRNA (vedi Figura 6)., I virus a RNA a singolo filamento come l’HIV trasportano un enzima speciale chiamato trascrittasi inversa all’interno del capside che sintetizza una copia complementare di ssDNA (cDNA) utilizzando il genoma +ssRNA come modello. L’ssDNA viene quindi trasformato in dsDNA, che può integrarsi nel cromosoma ospite e diventare una parte permanente dell’ospite. Il genoma virale integrato è chiamato provirus. Il virus ora può rimanere nell’ospite per lungo tempo per stabilire un’infezione cronica. Lo stadio di provirus è simile allo stadio di prophage in un’infezione batterica durante il ciclo lisogenico., Tuttavia, a differenza di prophage, il provirus non subisce l’escissione dopo la giunzione nel genoma.

Figura 6. Clicca per ingrandire l’immagine. L’HIV, un retrovirus icosaedrico avvolto, si attacca a un recettore di superficie cellulare di una cellula immunitaria e si fonde con la membrana cellulare. I contenuti virali vengono rilasciati nella cellula, dove gli enzimi virali convertono il genoma RNA a filamento singolo in DNA e lo incorporano nel genoma ospite., (credito: modifica del lavoro di NIAID, NIH)
Pensaci
- La RNA polimerasi RNA-dipendente è costituita da un gene virale o da un gene ospite?
Infezioni persistenti
L’infezione persistente si verifica quando un virus non viene completamente eliminato dal sistema dell’ospite ma rimane in determinati tessuti o organi della persona infetta. Il virus può rimanere in silenzio o subire un’infezione produttiva senza danneggiare seriamente o uccidere l’ospite., I meccanismi di infezione persistente possono comportare la regolazione delle espressioni geniche virali o dell’ospite o l’alterazione della risposta immunitaria dell’ospite. Le due categorie primarie di infezioni persistenti sono l’infezione latente e l’infezione cronica. Esempi di virus che causano infezioni latenti includono il virus herpes simplex (herpes orale e genitale), il virus varicella-zoster (varicella e herpes zoster) e il virus Epstein-Barr (mononucleosi). Il virus dell’epatite C e l’HIV sono due esempi di virus che causano infezioni croniche a lungo termine.,
Infezione latente
Non tutti i virus animali sono replicati dal ciclo litico. Ci sono virus che sono in grado di rimanere nascosti o dormienti all’interno della cella in un processo chiamato latenza. Questi tipi di virus sono noti come virus latenti e possono causare infezioni latenti. I virus in grado di latenza possono inizialmente causare un’infezione acuta prima di diventare dormienti.
Ad esempio, il virus varicella-zoster infetta molte cellule in tutto il corpo e causa la varicella, caratterizzata da un’eruzione di vesciche che coprono la pelle., Circa 10-12 giorni dopo l’infezione, la malattia si risolve e il virus diventa dormiente, vivendo all’interno dei gangli delle cellule nervose per anni. Durante questo periodo, il virus non uccide le cellule nervose o continua a replicarsi. Non è chiaro il motivo per cui il virus smette di replicarsi all’interno delle cellule nervose ed esprime poche proteine virali ma, in alcuni casi, in genere dopo molti anni di dormienza, il virus viene riattivato e causa una nuova malattia chiamata herpes zoster (Figura 7)., Mentre la varicella colpisce molte aree in tutto il corpo, l’herpes zoster è una malattia specifica delle cellule nervose che emerge dai gangli in cui il virus era dormiente.

Figura 7. (a) Varicella-zoster, il virus che causa la varicella, ha un capside icosaedrico avvolto visibile in questo micrografo elettronico di trasmissione. Il suo genoma del DNA a doppio filamento viene incorporato nel DNA ospite. (b) Dopo un periodo di latenza, il virus può riattivarsi sotto forma di herpes zoster, di solito manifestandosi come un’eruzione dolorosa e localizzata su un lato del corpo., (credito a: modifica del lavoro di Roskine Palmer e B. G. Partin—scala-bar dati da Matt Russell; credito b: modifica del lavoro di Rosmarie Voegtli)
I virus latenti possono rimanere dormienti da esistenti come molecole del genoma virale circolare al di fuori del cromosoma ospite. Altri diventano provirus integrandosi nel genoma ospite. Durante la dormienza, i virus non causano alcun sintomo di malattia e possono essere difficili da rilevare. Un paziente può essere inconsapevole di essere portatore del virus a meno che non sia stato eseguito un test diagnostico virale.,
Infezione cronica
Un’infezione cronica è una malattia con sintomi ricorrenti o persistenti per lungo tempo. Alcune infezioni virali possono essere croniche se il corpo non è in grado di eliminare il virus. L’HIV è un esempio di un virus che produce un’infezione cronica, spesso dopo un lungo periodo di latenza. Una volta che una persona viene infettata dall’HIV, il virus può essere rilevato nei tessuti in modo continuo, ma i pazienti non trattati spesso non presentano sintomi per anni., Tuttavia, il virus mantiene la persistenza cronica attraverso diversi meccanismi che interferiscono con la funzione immunitaria, compresa la prevenzione dell’espressione di antigeni virali sulla superficie delle cellule infette, alterando le cellule immunitarie stesse, limitando l’espressione dei geni virali e cambiando rapidamente gli antigeni virali attraverso la mutazione. Alla fine, il danno al sistema immunitario provoca la progressione della malattia che porta alla sindrome da immunodeficienza acquisita (AIDS)., I vari meccanismi che l’HIV utilizza per evitare di essere eliminato dal sistema immunitario sono utilizzati anche da altri virus infettivi cronici, incluso il virus dell’epatite C.
Pensaci
- In quali due modi un virus può riuscire a mantenere un’infezione persistente?
Ciclo di vita dei virus con ospiti vegetali
I virus vegetali sono più simili ai virus animali che ai batteriofagi. I virus delle piante possono essere avvolti o non avvolti. Come molti virus animali, i virus vegetali possono avere un genoma del DNA o del RNA ed essere singolo incagliato o doppio incagliato., Tuttavia, la maggior parte dei virus vegetali non ha un genoma del DNA; la maggior parte ha un genoma +ssRNA, che agisce come RNA messaggero (mRNA). Solo una minoranza di virus vegetali ha altri tipi di genomi.
I virus delle piante possono avere una gamma di host ristretta o ampia. Ad esempio, il virus citrus tristeza infetta solo poche piante del genere Citrus, mentre il virus del mosaico del cetriolo infetta migliaia di piante di varie famiglie di piante. La maggior parte dei virus vegetali sono trasmessi dal contatto tra le piante, o da funghi, nematodi, insetti o altri artropodi che agiscono come vettori meccanici., Tuttavia, alcuni virus possono essere trasferiti solo da un tipo specifico di vettore di insetti; ad esempio, un particolare virus potrebbe essere trasmesso dagli afidi ma non dalle mosche bianche. In alcuni casi, i virus possono anche entrare in piante sane attraverso ferite, come potrebbe verificarsi a causa di potatura o danni meteorologici.
I virus che infettano le piante sono considerati parassiti biotrofici, il che significa che possono stabilire un’infezione senza uccidere l’ospite, simile a quanto osservato nei cicli di vita lisogenici dei batteriofagi. L’infezione virale può essere asintomatica (latente) o può portare alla morte cellulare (infezione litica)., Il ciclo di vita inizia con la penetrazione del virus nella cellula ospite. Successivamente, il virus non è rivestito all’interno del citoplasma della cellula quando viene rimosso il capside. A seconda del tipo di acido nucleico, i componenti cellulari vengono utilizzati per replicare il genoma virale e sintetizzare le proteine virali per l’assemblaggio di nuovi virioni. Per stabilire un’infezione sistemica, il virus deve entrare in una parte del sistema vascolare della pianta, come il floema. Il tempo necessario per l’infezione sistemica può variare da pochi giorni a poche settimane a seconda del virus, delle specie vegetali e delle condizioni ambientali., Il ciclo di vita del virus è completo quando viene trasmesso da una pianta infetta a una pianta sana.
Pensaci
- Qual è la struttura e il genoma di un tipico virus vegetale?
Curva di crescita virale
A differenza della curva di crescita per una popolazione batterica, la curva di crescita per una popolazione virale nel suo ciclo di vita non segue una curva sigmoidale. Durante la fase iniziale, un inoculo di virus causa l’infezione. Nella fase di eclissi, i virus si legano e penetrano nelle cellule senza virioni rilevati nel mezzo., La principale differenza che appare successivamente nella curva di crescita virale rispetto a una curva di crescita batterica si verifica quando i virioni vengono rilasciati dalla cellula ospite lisata allo stesso tempo. Tale evento è chiamato scoppio e il numero di virioni per batterio rilasciato è descritto come la dimensione dello scoppio. In una curva di moltiplicazione a un passo per il batteriofago, le cellule ospiti si lisano, rilasciando molte particelle virali nel mezzo, il che porta ad un aumento molto ripido del titolo virale (il numero di virioni per unità di volume)., Se non rimangono cellule ospiti vitali, le particelle virali iniziano a degradarsi durante il declino della coltura (vedi Figura 8).
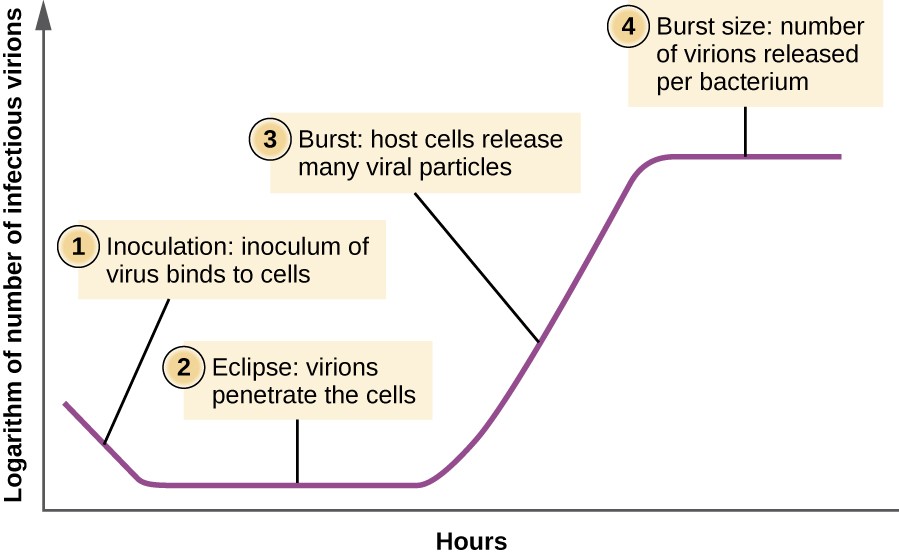
Figura 8. La curva di moltiplicazione a un passo per una popolazione di batteriofagi segue tre fasi: 1) inoculazione, durante la quale i virioni si attaccano alle cellule ospiti; 2) eclissi, durante la quale si verifica l’ingresso del genoma virale; e 3) scoppio, quando un numero sufficiente di nuovi virioni viene prodotto ed emerge dalla cellula ospite. La dimensione dello scoppio è il numero massimo di virioni prodotti per batterio.,
Pensaci
- Quale aspetto del ciclo di vita di un virus porta all’improvviso aumento della curva di crescita?
Trattamenti non registrati
Ebola è incurabile e mortale. L’epidemia in Africa occidentale nel 2014 è stata senza precedenti, sminuendo altre epidemie umane di Ebola nel livello di mortalità. Di 24.666 casi sospetti o confermati segnalati, 10.179 persone sono morte.
Non sono disponibili trattamenti o vaccini approvati per Ebola., Mentre alcuni farmaci hanno mostrato potenziale in studi di laboratorio e modelli animali, non sono stati testati sugli esseri umani per la sicurezza e l’efficacia. Non solo sono questi farmaci non testati o non registrati, ma sono anche a scarseggiare.
Data la grande sofferenza e gli alti tassi di mortalità, è giusto chiedersi se i farmaci non registrati e non testati siano meglio di nessuno. Tali farmaci dovrebbero essere dispensati e, in caso affermativo, chi dovrebbe riceverli, alla luce delle loro scorte estremamente limitate? È etico trattare farmaci non testati su pazienti con Ebola?, D’altra parte, è etico trattenere farmaci potenzialmente salvavita dai pazienti morenti? O forse i farmaci dovrebbero essere riservati agli operatori sanitari che lavorano per contenere la malattia?
Nell’agosto 2014, due operatori umanitari statunitensi infetti e un prete spagnolo sono stati trattati con ZMapp, un farmaco non registrato che era stato testato nelle scimmie ma non negli esseri umani. I due operatori umanitari americani si sono ripresi, ma il prete è morto. Più tardi quel mese, l’OMS ha pubblicato un rapporto sull’etica del trattamento dei pazienti con il farmaco., Dal momento che l’Ebola è spesso fatale, il gruppo ha motivato che è etico dare i farmaci non registrati e immorale trattenerli per problemi di sicurezza. Questa situazione è un esempio di” uso compassionevole ” al di fuori del consolidato sistema di regolamentazione e governance delle terapie.
Ebola negli Stati Uniti
Il 24 settembre 2014, Thomas Eric Duncan è arrivato al Texas Health Presbyterian Hospital di Dallas lamentando febbre, mal di testa, vomito e diarrea—sintomi comunemente osservati nei pazienti con raffreddore o influenza., Dopo l’esame, un medico del pronto soccorso gli diagnosticò sinusite, prescrisse alcuni antibiotici e lo mandò a casa. Due giorni dopo, Duncan tornò in ospedale in ambulanza. Le sue condizioni si erano deteriorate e ulteriori esami del sangue hanno confermato che è stato infettato dal virus Ebola.
Ulteriori indagini hanno rivelato che Duncan era appena tornato dalla Liberia, uno dei paesi nel bel mezzo di una grave epidemia di Ebola., Il 15 settembre, nove giorni prima di presentarsi all’ospedale di Dallas, Duncan aveva aiutato a trasportare un vicino colpito da Ebola in un ospedale in Liberia. L’ospedale ha continuato a curare Duncan, ma è morto diversi giorni dopo essere stato ricoverato.

Figura 9. I ricercatori che lavorano con il virus Ebola utilizzano strati di difese contro le infezioni accidentali, inclusi indumenti protettivi, sistemi respiratori e armadi a pressione negativa per il lavoro su banco. (credit: modification of work di Randal J., Schoepp)
La cronologia del caso Duncan è indicativa del ciclo di vita del virus Ebola. Il tempo di incubazione per Ebola varia da 2 giorni a 21 giorni. Passarono nove giorni tra l’esposizione di Duncan all’infezione da virus e la comparsa dei suoi sintomi. Ciò corrisponde, in parte, al periodo di eclissi nella crescita della popolazione del virus. Durante la fase di eclissi, Duncan non sarebbe stato in grado di trasmettere la malattia ad altri. Tuttavia, una volta che un individuo infetto inizia a mostrare sintomi, la malattia diventa molto contagiosa., Il virus Ebola viene trasmesso attraverso il contatto diretto con goccioline di fluidi corporei come saliva, sangue e vomito. Duncan potrebbe plausibilmente aver trasmesso la malattia ad altri in qualsiasi momento dopo aver iniziato ad avere sintomi, presumibilmente qualche tempo prima del suo arrivo in ospedale a Dallas. Una volta che un ospedale si rende conto che un paziente come Duncan è infetto dal virus Ebola, il paziente viene immediatamente messo in quarantena e i funzionari della sanità pubblica iniziano una traccia posteriore per identificare tutti con i quali un paziente come Duncan potrebbe aver interagito durante il periodo in cui stava mostrando i sintomi.,
I funzionari della sanità pubblica sono stati in grado di rintracciare 10 individui ad alto rischio (membri della famiglia di Duncan) e 50 individui a basso rischio per monitorarli per i segni di infezione. Nessuno ha contratto la malattia. Tuttavia, una delle infermiere incaricate delle cure di Duncan si infettò. Questo, insieme alla diagnosi errata iniziale di Duncan, ha chiarito che gli ospedali statunitensi avevano bisogno di fornire ulteriore formazione al personale medico per prevenire una possibile epidemia di Ebola negli Stati Uniti.
- Quali tipi di formazione possono preparare gli operatori sanitari a contenere epidemie emergenti come l’epidemia di Ebola del 2014?,
- Qual è la differenza tra un agente patogeno contagioso e un agente patogeno infettivo?
Concetti chiave e riepilogo
- Molti virus colpiscono specifici host o tessuti. Alcuni possono avere più di un host.
- Molti virus seguono diverse fasi per infettare le cellule ospiti. Queste fasi includono attaccamento, penetrazione, svolgimento, biosintesi, maturazione e rilascio.
- I batteriofagi hanno un ciclo litico o lisogenico. Il ciclo litico porta alla morte dell’ospite, mentre il ciclo lisogenico porta all’integrazione del fago nel genoma dell’ospite.,
- I batteriofagi iniettano il DNA nella cellula ospite, mentre i virus animali entrano per endocitosi o fusione di membrana.
- I virus animali possono subire latenza, simile alla lisogenesi per un batteriofago.
- La maggior parte dei virus vegetali sono ssRNA a filamento positivo e possono subire latenza, infezione cronica o litica, come osservato per i virus animali.
- La curva di crescita delle popolazioni di batteriofagi è una curva di moltiplicazione a un passo e non una curva sigmoidale, rispetto alla curva di crescita batterica.,
- I batteriofagi trasferiscono informazioni genetiche tra gli host utilizzando la trasduzione generalizzata o specializzata.
Scelta multipla
Quale delle seguenti porta alla distruzione delle cellule ospiti?
- lisogeni ciclo
- ciclo litico
- profago
- fago temperato
Un virus ottiene il suo involucro durante quale delle seguenti fasi?,
- attachment
- penetration
- assembly
- release
Quale dei seguenti componenti viene portato in una cellula dall’HIV?
- una DNA-polimerasi DNA dipendente
- RNA polimerasi
- ribosoma
- trascrittasi inversa
Un virus a RNA a filamento positivo:
- deve prima essere convertito in un mRNA prima che possa essere tradotto.
- può essere utilizzato direttamente per tradurre le proteine virali.
- sarà degradato dagli enzimi ospiti.
- non è riconosciuto dai ribosomi dell’ospite.
Qual è il nome per il trasferimento di informazioni genetiche da un batterio a un altro batterio da parte di un fago?,
- trasduzione
- penetrazione
- escissione
- traduzione
Pensaci
- Spiega brevemente la differenza tra il meccanismo di ingresso di un batteriofago T-even e un virus animale.
- Discutere la differenza tra trasduzione generalizzata e specializzata.,
- Distinguere tra cicli litici e lisogenici.
- I batteriofagi hanno cicli litici e lisogenici. Discutere i vantaggi e gli svantaggi per il fago.
- In che modo la trascrittasi inversa aiuta un retrovirus a stabilire un’infezione cronica?
- Discutere alcuni metodi con cui i virus vegetali vengono trasmessi da una pianta malata a una sana.
- Etichettare i cinque stadi di un’infezione da batteriofagi nella figura:
