embryonale homologieën
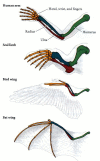
een van de belangrijkste onderscheidingen gemaakt door de evolutionaire embryologen was het verschil tussen analogie en homologie. Beide termen verwijzen naar structuren die vergelijkbaar lijken te zijn. Homologe structuren zijn die organen waarvan de onderliggende gelijkenis voortkomt uit hun zijn afgeleid van een gemeenschappelijke voorouderlijke structuur. Bijvoorbeeld, de vleugel van een vogel en de voorpoot van een mens zijn homoloog. Bovendien zijn hun respectieve delen homoloog (figuur 1.13)., Analoge structuren zijn structuren waarvan de gelijkenis voortkomt uit het uitvoeren van een soortgelijke functie, in plaats van uit een gemeenschappelijke voorouder. Daarom zijn bijvoorbeeld de vleugel van een vlinder en de vleugel van een vogel analoog. De twee soorten vleugels hebben een gemeenschappelijke functie (en worden daarom beide vleugels genoemd), maar de vogelvleugel en de insectenvleugel zijn niet ontstaan uit een oorspronkelijke voorouderlijke structuur die door de evolutie werd gewijzigd in vogelvleugels en vlindervleugels.
figuur 1.,13
homologieën van structuur tussen een menselijke arm, een voorpoot van een zeehond, een vogelvleugel en een vleermuisvleugel; homologe draagstructuren zijn in dezelfde kleur weergegeven. Alle vier zijn homoloog als voorpoten en zijn afgeleid van een gemeenschappelijke tetrapod voorouder. Huisvestingen (meer…)
homologieën moeten zorgvuldig worden gemaakt en moeten altijd verwijzen naar het niveau van organisatie dat wordt vergeleken. Zo zijn de vogelvleugel en de vleermuisvleugel homoloog als voorpoten, maar niet als vleugels., Met andere woorden, ze delen een gemeenschappelijke onderliggende structuur van voorpoten omdat vogels en zoogdieren een gemeenschappelijke afkomst hebben. De vogelvleugel ontwikkelde zich echter onafhankelijk van de vleermuisvleugel. Vleermuizen stammen af van een lange lijn van niet-vleugelige zoogdieren, en de structuur van de vleermuisvleugel is duidelijk verschillend van die van een vogelvleugel.een van de meest gevierde gevallen van embryonale homologie is dat van het kieuwkraakbeen van vissen, de reptielenkaak en het middenoor van zoogdieren (beoordeeld in Gould 1990)., Eerst werden de kieuwbogen van kaakloze (agnathan) vissen aangepast om de kaak van de jawed vissen te vormen. In de kaakloze vissen, een reeks kieuwen geopend achter de kaakloze mond. Toen de kieuwspleten werden ondersteund door kraakbeenelementen, omringde de eerste set van deze kieuwsteunen de mond om de kaak te vormen. Er is voldoende bewijs dat kaken gemodificeerde kieuwsteunen zijn. Ten eerste, beide sets botten zijn gemaakt van neurale kam cellen. (De meeste andere botten komen uit mesodermale Weefsel.) Ten tweede, beide structuren vormen van de bovenste en onderste bars die naar voren buigen en scharnierend in het midden., Ten derde lijkt de kaakspier homoloog aan de oorspronkelijke kieuwsteunspier. De gewervelde kaak lijkt dus homoloog te zijn aan de kieuwbogen van kaakloze vissen.
maar het verhaal eindigt hier niet. Het bovenste gedeelte van de tweede embryonale boog ter ondersteuning van de kieuw werd het hyomandibulaire bot van jawed vissen. Dit element ondersteunt de schedel en verbindt de kaak met de schedel (figuur 1.14 A). Toen gewervelde dieren op het land kwamen, hadden ze een nieuw probleem: hoe te horen in een medium zo dun als lucht., Het hyomandibulaire bot toevallig in de buurt van de otic (oor) capsule, en benige materiaal is uitstekend voor het overbrengen van geluid. Zo begon het hyomandibulaire bot van de eerste amfibieën, terwijl het nog steeds functioneerde als een schedelbrace, ook te functioneren als een geluidsomvormer (Clack 1989). Toen de gewervelde dieren hun beweging, kaakstructuur en houding veranderden, werd de schedel stevig aan de rest van de schedel gehecht en had de hyomandibulaire brace niet nodig. Het hyomandibulaire bot lijkt dan gespecialiseerd te zijn geworden in de stapes bot van het middenoor., Wat de secundaire functie van dit bot was, werd zijn primaire functie.
de oorspronkelijke kaakbeenderen veranderden ook. De eerste embryonale boog genereert het kaakapparaat. Bij amfibieën, reptielen en vogels vormt het achterste deel van dit kraakbeen het quadrate bot van de bovenkaak en het articulaire bot van de onderkaak. Deze botten verbinden zich met elkaar en zijn verantwoordelijk voor het articuleren van de boven-en onderkaak. Bij zoogdieren gebeurt deze articulatie echter op een ander gebied( de dentaire en squamosale botten), waardoor deze benige elementen worden “vrijgemaakt” om nieuwe functies te verwerven., Het quadrate bot van de Reptiel bovenkaak evolueerde tot het zoogdierincusbot van het middenoor, en het articulaire bot van de onderkaak van het reptiel is onze malleus geworden. Dit laatste proces werd voor het eerst beschreven door Reichert in 1837, toen hij in het varkensembryo observeerde dat de onderkaak (kaakbeen) verbreedt aan de zijkant van Meckels kraakbeen, terwijl het achterste deel van Meckels kraakbeen verbreedt, loskomt van de rest van het kraakbeen, en het middenoor binnendringt om de malleus te worden (figuur 1.14 B,C)., Zo zijn de middenoorbotten van het zoogdier homoloog aan de achterste onderkaak van het reptiel en aan de kieuwbogen van agnathan vissen. In hoofdstuk 22 wordt nader ingegaan op Meer recente informatie over de relatie tussen ontwikkeling en evolutie.