embryonala homologier
en av de viktigaste skillnaderna från de evolutionära embryologerna var skillnaden mellan analogi och homologi. Båda termerna hänvisar till strukturer som verkar vara likartade. Homologa strukturer är de organ vars underliggande likhet härrör från att de härrör från en gemensam förfaderstruktur. Till exempel är en fågelns vinge och en människas framben homologa. Dessutom är deras respektive delar homologa (figur 1.13)., Analoga strukturer är de vars likhet kommer från att de utför en liknande funktion, snarare än att de härrör från en gemensam förfader. Därför är till exempel vingen av en fjäril och vingen av en fågel analog. De två typerna av vingar delar en gemensam funktion (och kallas därför båda vingarna), men fågelvingen och insektsvingen uppstod inte från en original förfaderstruktur som blev modifierad genom utveckling till fågelvingar och fjärilsvingar.
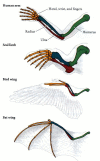
Figur 1.,13
homologier av struktur bland en mänsklig arm, en tätning forelimb, en fågelvinge och en fladdermus vinge; homologa stödstrukturer visas i samma färg. Alla fyra är homologa som förben och härleddes från en vanlig tetrapod förfader. Anpassningarna (mer…)
Homologier måste göras noggrant och måste alltid hänvisa till organisationsnivån som jämförs. Till exempel är fågelvingen och batvingen homologa som förben, men inte som vingar., Med andra ord delar de en gemensam underliggande struktur av forelimbben eftersom fåglar och däggdjur delar ett gemensamt anor. Fågelvingen utvecklades dock oberoende av fladdermusvingen. Fladdermöss härstammar från en lång rad nonwinged däggdjur, och strukturen av bat vingen skiljer sig markant från en fågel vinge.
ett av de mest berömda fallen av embryonal homologi är det av fiskgillbrusk, reptilkäken och däggdjursörat (granskat i Gould 1990)., Först blev gillbågarna av jawless (agnathan) fiskar modifierade för att bilda käken av de jawed Fiskarna. I de jawless Fiskarna öppnades en serie gälar bakom den jawless munnen. När gillslitsarna stöddes av broskiga element, omgav den första uppsättningen av dessa gillstöd munnen för att bilda käften. Det finns gott om bevis för att käftarna är modifierade gill stöder. För det första är båda dessa uppsättningar ben gjorda av neurala kamceller. (De flesta andra ben kommer från mesodermal vävnad.) För det andra bildar båda strukturerna från övre och nedre stänger som böjer sig framåt och är gångjärn i mitten., För det tredje verkar käftmuskulaturen vara homolog till den ursprungliga gill-stödmuskulaturen. Således verkar vertebratkäken vara homolog till gillbågarna av jawless fiskar.
men historien slutar inte här. Den övre delen av den andra embryonala bågen som stöder gillen blev det hyomandibulära benet av jawed fiskar. Detta element stöder skallen och länkar käften till kraniet (figur 1.14 a). När ryggradsdjur kom upp på land hade de ett nytt problem: hur man hör i ett medium så tunt som luft., Det hyomandibulära benet råkar vara nära Otic (ear) kapseln, och benigt material är utmärkt för överföring av ljud. Således, medan den fortfarande fungerar som en kranialstag, började de första amfibiernas hyomandibulära ben också fungera som en ljudgivare (Clack 1989). När de markbundna ryggkotorna förändrade deras rörelse, käftstruktur och hållning blev kraniet fast fastsatt på resten av skallen och behövde inte den hyomandibulära stag. Det hyomandibulära benet verkar då ha blivit specialiserat på mellanörets stapeben., Vad som hade varit Detta Ben sekundära funktion blev dess primära funktion.
de ursprungliga käftbenen ändrades också. Den första embryonala bågen genererar käftapparaten. I amfibier, reptiler och fåglar bildar den bakre delen av detta brosk quadratbenet i överkäken och ledbenet i underkäken. Dessa ben ansluter till varandra och är ansvariga för att artikulera de övre och nedre käftarna. Men hos däggdjur sker denna artikulation i en annan region (de kort-och squamosala benen), vilket ”frigör” dessa beniga element för att förvärva nya funktioner., Kvadratbenet i reptilens överkäke utvecklades till det däggdjursincus benet i mellanörat, och det artikulära benet i reptilens underkäke har blivit vår malleus. Denna senare process beskrevs först av Reichert 1837, när han observerade i grisembryot att mandibeln (käftbenet) förbenar på sidan av Meckels brosk, medan den bakre delen av Meckels brosk förbenar, lossnar från resten av brosket och går in i mellanörat för att bli malleus (figur 1.14 B, C)., Således är däggdjurets mellanöraben homologa till reptilens bakre underkäke och till agnathans Fiskarnas gillbågar. Kapitel 22 kommer att innehålla mer aktuell information om utvecklingsförhållandet till utvecklingen.