Obrázek 1. Kanonická a nekanonická signalizační dráha NF-kB
Co je NF-kB?
NF-kB je krátký název jaderného faktoru kappa-light-chain-enhancer aktivovaných B buněk. Není to jediný protein, ale malá rodina indukovatelných transkripčních faktorů, které hrají důležitou roli téměř ve všech savčích buňkách. Řídí transkripci DNA, produkci cytokinů, přežití buněk a další důležité buněčné události, zejména hrají klíčovou roli při regulaci imunitní odpovědi na infekci.
NF-kB molekulární jsou obvykle dimery., Typická struktura NF-kB je P50-P65 dimer (NF-kB1/RelA). Tvorba dimeru je nezbytná pro vazbu DNA, dva monomery NF-kB se váží na DNA jako dimer. N-terminální oblasti dimeru jsou zodpovědné za specifický kontakt DNA. C-terminální oblasti jsou obvykle vysoce konzervované, oni jsou zodpovědní za dimerizace a nespecifické DNA fosfát kontakt. Celá molekula NF-kB je stejně jako kleště svěráku na řetězci DNA a funguje jako transkripční faktor (obrázek 2).
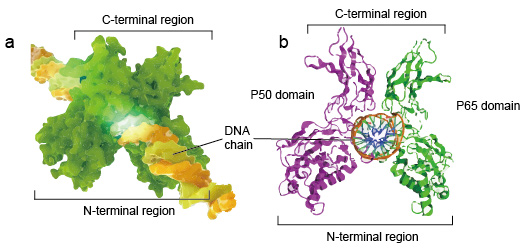
Obrázek 2. Struktura vazby NF-kB proteinového dimeru s řetězcem DNA.,
rodina NF-kB
protože členové rodiny NF-kB sdílejí strukturální homologii s retrovirovým onkoproteinem v-Rel. takže také nazývali proteiny NF-kB / Rel. NF-kB transkripční faktor rodiny u savců se skládá z 5 proteinů: p65/Rel, RelB, c-Rel, p105/p50 (NF-kB1), a p100/p52 (NF-kB2). Těchto 5 proteinů se navzájem spojuje a tvoří zřetelné transkripčně aktivní homo / heterodimerní komplexy (obrázek 3). Všichni sdílejí společnou konzervovanou Rel homology doménu (RHD)o délce 300 aa., Tato doména RHD má více funkcí, jako je dimerizace, vazba DNA, interakce s IkBs a jaderná translokace. Ačkoli členové rodiny proteinů NF-kB mohou tvořit až 15 různých dimerů, ale mnoho z nich dosud neprokázalo. Nejhojnější formou NF-kB dimeru je P50 / P65 heterodimer, který byl financován téměř ve všech typech buněk. Protože pouze P65 / Rel, RelB a c-Rel mají karboxy-terminální transaktivační domény (TAD), lze rodinu proteinů NF-kB dále rozdělit do dvou skupin., p50 a p52 jsou generovány zpracováním prekurzorových molekul p105 a p100. Proto ne všechny kombinace Rel dimerů jsou transkripčně aktivní.
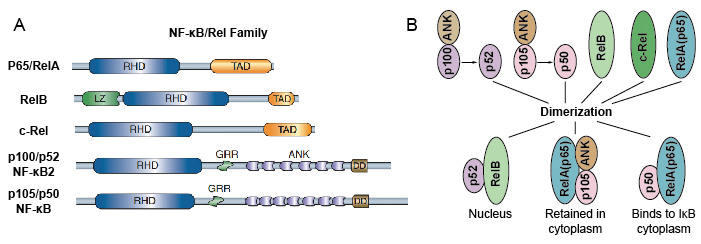
obrázek 3. Protein 5 a jejich homo / heterodimerní komplexy rodiny NF-kB.
signalizační dráha NF-kB
stmívače proteinů NF-kB jako faktor nukleární transkripce musí migrovat do jádra v kombinaci s DNA, aby měly funkci. Ve většině typů normálních buněk v klidovém stavu byla NF-KB neaktivní a udržovala se v cytoplazmě., Oni vazbou na specifické inhibitory nazývá IK-B protein, který se může vázat na Rel homology domain (RHD) NF-kB a v rozporu s jeho jaderné lokalizační sekvence (NLS) funkce. Tyto inhibitor proteinů, které zahrnují IkBa, IkBb a IkBg, obsahují 6-7 ankyrin opakuje, že zprostředkovávají vazbu na RHD. Tyto repetice jsou také přítomný v C-terminální poloviny NF-kB2/p100 a NF-kB1/p105 prekurzory, které rovněž fungují jako IkBs a udržet si své partnery, Rel proteinů v cytoplazmě., Aby bylo možné aktivovat molekulu NF-kB, musí buňky nejprve oddělit protein NF-kB od svých inhibitorů. Tam jsou dvě hlavní signální dráhy vedou k IK-B protein inhibitor disociace od NF-kB stmívače a nechat translokace NF-kB dimery z cytoplazmy do jádra.
1., Canonical/Klasické Kaskády
Canonical nebo klasické kaskády signalizace od povrchu buňky receptor pro-zánětlivých cytokinů a patogen-associated molecular patterns (PAMPs), jako jsou tumor nekrotizující faktor receptor (TNFR), toll-like receptorů (TLR) a T/B buněčný receptor. Tyto receptory vazba s jejich ligand molekuly a přenos signálu přes buněčnou membránu, způsobit aktivaci IkB kinázy (IKK) komplex. Nejběžnější formou tohoto komplexu je heterodimer katalytických podjednotek IKKa a IKKß a regulační podjednotka IKKy., Jednotka IKKy také nazývala NEMO pro NF-kB základní modulátor. Aktivuje IKK komplex, převážně jednající prostřednictvím IKKß v IKKy-závislým způsobem, katalyzuje fosforylaci IkBs (v místech ekvivalentní Ser32 a Ser36 z IkBa), polyubiquitination (v místech ekvivalentní Lys21 a Lys22 z IkBa) a následné degradaci v proteazomu 26S. Uvolněné dimery NF-kB (nejčastěji dimer P50–RelA), translokují se do jádra, váží DNA a aktivují transkripci genu down-stream.
2., Nekanonická / alternativní dráha Kaskáda
další aktivační dráha NF-kB nazývaná ne-kanonická nebo alternativní cesta. Tato cesta je nezávislá na IKKß a IKKy, ale místo toho závisí na stmívači IKKa. Signalizační přenos do cytoplazmy přes LT-β nebo BAFF receptor. Fosfonát a NIK protein, NIK protein pak fosfonovat ikka homodimmery. Cílem Homodimerů IKKa v této cestě je NF-KB2/p100, který je fosforylován na dvou místech C-terminálů., Fosforylace těchto míst je nezbytná pro zpracování p100 na p52, což je také závislé na polyubiquitinaci a proteazomální degradaci. Nicméně, místo toho, vedoucí k úplné p100 rozkladu, jak je vidět s IkBs, fosforylaci závislé ubikvitinace, z p100 výsledky pouze v degradaci jeho inhibiční C-terminální polovina dílů. Jakmile je polovina C-terminálu degradována, uvolní se n-koncová část NF-kB (P52 polypeptid, který obsahuje RHD)., Jako RHD z p100 je nejčastěji spojován s RelB, aktivace této „alternativní“ cestou, výsledky v jaderné translokaci p52–RelB dimery. Dimer se nakonec váže na DNA a aktivuje transkripci genu down-stream.
3. Cestou nařízení,
Ubiquitin je 76 aminokyselin proteinu, který je vysoce konzervovaným a všude a vyjádřené ve všech eukaryot z kvasinek na člověka. Karboxylová kyselina v C-terminálním glycinu ubiquitinu může být kovalentně připojena k Epsilon Aminu lysinu na jiném proteinu prostřednictvím isopeptidové vazby. Tento proces se nazývá „ubiquitinace“., Navíc, C-terminální ocas ubiquitin může být přímo připojen na N-terminální methionin jiného ubiquitin tvoří lineární polyubiquitin řetězce. Ubiquitinace hraje zásadní roli při regulaci NF-kB drah. V nestimulovaných buňkách se NF-kB váže na inhibiční proteiny rodiny kB (IkB) a je sekvestrován v cytoplazmě., Po stimulaci, IkB je fosforylován prostřednictvím IkB kinázy (IKK) komplexní, Fosforylovaný IkB je následně ubiquitní a degradován v proteazomu 26S, což umožňuje NF-kB přemisťovat do jádra, kde reguluje expresi velkého počtu genů. Jako regulační podjednotky IKK komplex, NEMO je navržen ke klíčovým faktorem pro transdukce ubikvitinace signál aktivace IKK. Několik Dabuje funkce jako klíčové negativní regulátory IKK umožňující přísnou kontrolu NF-kB aktivace., Jeden z nejlepších studoval Dabuje je A20, bylo navrženo, že A20 potlačit hyperaktivace NFkB tím, že deubiquitinating K63-spojené polyubiquitin připojené k RIP1 prostřednictvím N-terminální OTU domény, stejně jako podpora K48-spojené polyubiquitination RIP1 pro proteasomal degradace.
4. Následné signalizační
NF-kB působí jako ústřední mediátor imunitních a zánětlivých reakcí, a je zapojen do stresové reakce a regulace buněčné proliferace a apoptózy. Příslušné cílové geny NF-kB umožňují organismu účinně reagovat na tyto změny prostředí., Klasická cesta NF-kB je aktivována různými zánětlivými signály, což vede k koordinované expresi více zánětlivých a vrozených imunitních genů. Na prozánětlivých cytokinů IL-1b a TNF-a aktivaci NF-kB, a jejich exprese je indukována v odpovědi na NF-kB aktivace, čímž tvoří zesilování posuv smyčky. Alternativní cesta pro NF-kB výsledky v jaderné translokaci p52–RelB dimery, je přísně závislé na IKKa homodimers a je aktivován lt br, BAFF a CD40L do NIK., Mnoho údajů silně naznačuje, že alternativní cesta hraje ústřední roli při expresi genů podílejících se na vývoji a udržování sekundárních lymfoidních orgánů. Transkripční faktory NF-kB rodiny kontrolují expresi velkého počtu cílových genů v reakci na změny v prostředí, čímž pomáhá organizovat zánětlivé a imunitní odpovědi.
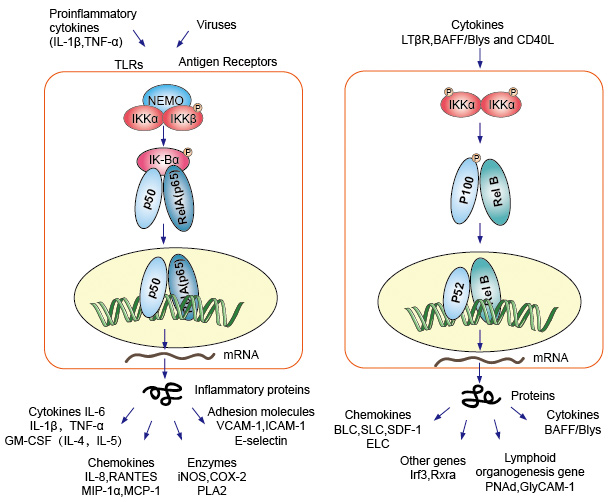
obrázek 4. Signalizace NF-kB signalizační dráhy.
5. Vztah s nemocemi.
- zánětlivý základ metabolických onemocnění.,
signalizační dráha IKK / NF-kB je klíčem k propojení metabolismu, zánětu a účinku inzulínu. Většina metabolický stres signály, což vede k inzulínové rezistence nebo pankreatických β buněk dysfunkce, ať už vyvolaný intracelulární nebo extracelulární podněty, sbíhají na NF-kB aktivace kinázy IKKb a další významné zánětlivé kinázy JNK-mitogen aktivované proteinkinázy (MAPK).
- NF-kB a glykolýza.
glykolýza může generovat ATP ve větším množství a rychleji než oxidační fosforylace., Glukóza je proto nezbytnou živinou pro rakovinu a normální proliferující buňky. NF-kB koordinuje mnoho signálů, které řídí aktivaci a proliferaci buněk během imunity, zánětu a onkogeneze.
- NF-kB a oxidační metabolismus.
NF-kB upravuje energetickou homeostázu a metabolickou adaptaci kontrolou rovnováhy mezi glykolýzou a dýcháním pro poskytování energie. Inhibice NF-kB / RelA různými prostředky snížila spotřebu kyslíku a způsobila přeprogramování na aerobní glykolýzu