Figur 1. De kanoniske og ikke-kanoniske NF-kB signaliserer sti
Hva er NF-kB?
NF-kB er et kort navn av Kjernefysiske Faktor kappa-lys-kjede-enhancer-aktiverte B-celler. Det er ikke et enkelt protein, men en liten familie av inducible transkripsjon faktorer som spiller en viktig rolle i nesten alle mammalske celler. Det kontroll-DNA transkripsjon, cytokin produksjon, celle overlevelse og andre viktige celle arrangementer, spesielt spille en viktig rolle i regulering av immunrespons mot infeksjon.
NF-kB molekylær er vanligvis dimers., En typisk struktur av NF-kB er P50-P65 dimer (NF-kB1/RelA). Den dimer dannelse er nødvendig for DNA-bindende, to NF-kB monomers binde seg til DNA som en dimer. N-terminal regioner av dimer er ansvarlig for bestemte DNA-kontakt. C-terminal regioner er vanligvis svært bevart, de er ansvarlig for dimerization og uspesifikke DNA-fosfat kontakt. Hele NF-kB molekylær er akkurat som en tang skrustikke på DNA-kjeden, og fungerer som en transkripsjonsfaktor (figur 2).
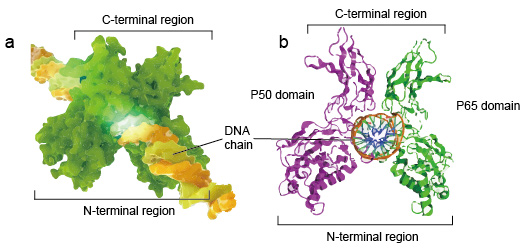
Figur 2. Strukturen av NF-kB protein dimer bindende med DNA-kjeden.,
NF-kB familie
Fordi NF-kB familiemedlemmer dele strukturell homologi med retroviral oncoprotein v-Rel. slik at de også kalles NF-kB/Rel Proteiner. NF-kB transkripsjonsfaktor familie i pattedyr består av 5 proteiner: p65/RelA, RelB, c-Rel, p105/p50 (NF-kB1), og p100 – /p52 (NF-kB2). Disse 5 proteiner omgås med hverandre og danner distinkte transcriptionally aktiv homo/heterodimeric komplekser (Figur 3). De deler alle en felles bevart Rel homologi domene (RHD) av 300 aa lengde., Disse RHD domenet har flere funksjoner som dimerization, DNA-bindende, samhandling med IkBs, og kjernefysiske translocation. Selv om NF-kB protein familiemedlemmer kan danne opp til 15 forskjellige dimers, men mange av dem har ennå ikke vist. De mest tallrike form av NF-kB-dimer er p50/p65 heterodimer som har vært fondet i nesten alle celletyper. Som bare P65/Rel, RelB og c-Rel har carboxy-terminal transactivation domener (TAD), NF-kB familie av proteiner kan videre deles inn i to grupper., p50 og p52 er generert ved behandling av visse molekyler p105 og p100 -, henholdsvis. Derfor er ikke alle kombinasjoner av Rel dimers er transcriptionally aktiv.
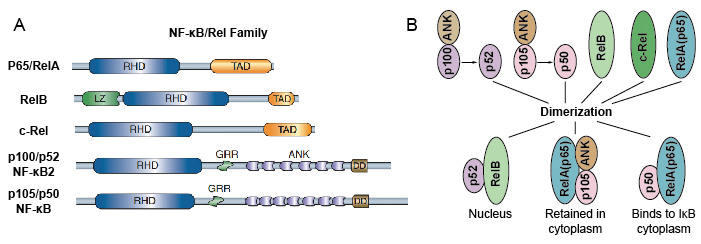
Figur 3. 5 protein og deres homo/heterodimeric komplekser av NF-kB familie.
NF-kB signaliserer sti
NF-kB protein dimmere som kjernefysiske transkripsjonsfaktor, de trenger å migrere til kjernen, kombinert med DNA-å ha funksjon. I de fleste typer av normale celler under hvilende tilstand, NF-KB var inaktive og beholde i cytoplasma., De binding til en bestemt hemmere kalt IK-B protein, noe som kan binde seg til Rel homologi domene (RHD) av NF-kB og forstyrre sin kjernefysiske lokalisering sekvens (NLS) – funksjonen. Disse hemmer proteiner, som inkluderer IkBa, IkBb og IkBg, inneholde 6-7 ankyrin gjentar at megle binding til RHD. Disse gjentar, er også til stede i C-terminal halvdelene av NF-kB2/p100-og NF-kB1/p105 forløpere, som også fungerer som IkBs og beholde sine partnere, Rel proteiner, i cytoplasma., For å aktivere NF-kB-molekyler, celler, først må du skille NF-kB protein fra deres inhibitors. Det er to store signaliserer veier fører til IK-B protein som hemmer dissosiasjon fra NF-kB dimmer og la translocation av NF-kB dimers fra cytoplasma inn i kjernen.
1., Kanonisk/Klassiske Cascade
Kanonisk eller klassiske cascade signalering fra celleoverflaten reseptor av pro-inflammatoriske cytokiner og patogen-associated molecular patterns (PAMPs) slik som tumor nekrose faktor reseptor (TNFR), toll-like receptor (TLR) og T/B-celle reseptor. Disse reseptorene bindende med sin ligand molekyler og overføre signalet over cellemembranen, føre til aktivering av IkB kinase (IKK) – komplekset. Den mest vanlige formen av dette komplekset, som består heterodimer av IKKa og IKKß katalytisk underenhetene og en IKKy regulatoriske subunit., Den IKKy enhet, også kalt NEMO for NF-kB viktig å begå. Den aktiverte IKK-komplekset, hovedsakelig handler gjennom IKKß i en IKKy-avhengig måte, catalyzes den phosphorylation av IkBs (på nettsteder som tilsvarer Ser32 og Ser36 av IkBa), polyubiquitination (på nettsteder som tilsvarer Lys21 og Lys22 av IkBa) og påfølgende degradering av 26S proteasome. Utgitt NF-kB dimers (vanligst på p50–RelA dimer), translocate til kjernen, binder DNA og aktiver ned-stream gentranskripsjon.
2., Ikke-kanoniske/Alternative veien Cascade
en Annen NF-kB aktivere sti kalles ikke-kanoniske eller alternative veien. Denne veien er uavhengig av IKKß og IKKy, men avhengig av IKKa dimmer i stedet. Signalene overføres til cytoplasma gjennom LT-β eller BAFF reseptor. Phosphonate en NIK protein, NIK protein så phosphonate det IKKa homodimmers. Målet for IKKa homodimers i denne veien er NF-kB2/p100 -, som er phosphorylated på to C-terminal nettsteder., Phosphorylation av disse områdene er avgjørende for p100-behandling til p52, som også er avhengig av polyubiquitination og proteasomal nedbrytning. Men i stedet for å ledende for å fullføre p100-degradering, som sett med IkBs, den phosphorylation-avhengige ubiquitination av p100-resultater bare i nedbrytning av sin hemmende C-terminal halvparten deler. Når C-terminal halvparten er degradert, N-terminal del av NF-kB (p52 polypeptid som inneholder RHD) er utgitt., Som RHD av p100-er som oftest forbundet med RelB, aktivering av denne ‘alternativ’ vei resultater i kjernefysiske translocation av p52–RelB dimers. Den dimer til slutt binder seg til DNA og aktiv down-stream gentranskripsjon.
3. Veien regulering
Ubiquitin er en 76-amino-acid protein som er godt bevart og ubiquitously til uttrykk i alle eukaryotes fra gjær til menneske. Den karboksylsyre i C-terminal glysin av ubiquitin kan være kovalent bundet til epsilon amine av lysin på en annen protein gjennom en isopeptide bond. Denne prosessen kalles «ubiquitination»., Videre, C-terminal hale av ubiquitin kan være direkte koblet til N-terminal metionin av en annen ubiquitin å danne lineær polyubiquitin kjede. Ubiquitination spiller en viktig rolle i reguleringen av NF-kB trasé. I unstimulated celler, NF-kB binder seg til hemmende proteiner av kB familie (IkB) og er trukket i cytoplasma., Ved stimulering, IkB er phosphorylated av IkB kinase (IKK) – kompleks, Phosphorylated IkB er senere ubiquitinated og degradert av 26S proteasome, slik at NF-kB til translocate til kjernen, hvor det regulerer uttrykk av en mengde gener. Som regel subunit av IKK-komplekset, NEMO har blitt foreslått for å være den viktigste faktoren for transducing ubiquitination signal til IKK aktivering. Flere Kaller funksjonen som tasten negative regulatorer av IKK å tillate en streng kontroll av NF-kB aktivering., En av de best studerte Kaller er A20, Det har blitt foreslått at A20 undertrykke hyperactivation av NFkB av deubiquitinating K63 knyttet polyubiquitin knyttet til RIP1 via N-terminal OTU domene, samt å fremme K48 knyttet polyubiquitination av RIP1 for proteasomal nedbrytning.
4. Nedstrøms signaliserer
NF-kB fungerer som en sentral formidler av immunsystemet og inflammatorisk respons, og er involvert i stressrespons og regulering av celleproliferasjon og apoptosis. De respektive NF-kB mål gener tillate organismen til å reagere effektivt på disse miljømessige endringer., Den klassiske NF-kB vei er aktivert av en rekke inflammatoriske signaler, noe som resulterer i koordinering uttrykk for flere inflammatorisk og medfødte immunsystemet gener. Den proinflammatory cytokiner som IL-1b og TNF-a aktiver NF-kB, og deres uttrykk er indusert i respons til NF-kB-aktivering, og dermed danner en forsterkende feed forward loop. Den alternative veien for NF-kB resultater i kjernefysiske translocation av p52–RelB dimers, er strengt avhengig av IKKa homodimers og aktiveres ved LTbR, BAFF og CD40L av NIK., Mange data antyder sterkt at den alternative veien spiller en sentral rolle i uttrykket av gener som er involvert i utvikling og vedlikehold av sekundære lymfoide organer. Transkripsjon faktorer av NF-kB-familien styre uttrykk for et stort antall mål-gener i respons til endringer i miljøet, og dermed bidra til å legge til rette for inflammatorisk og immunsystemet.
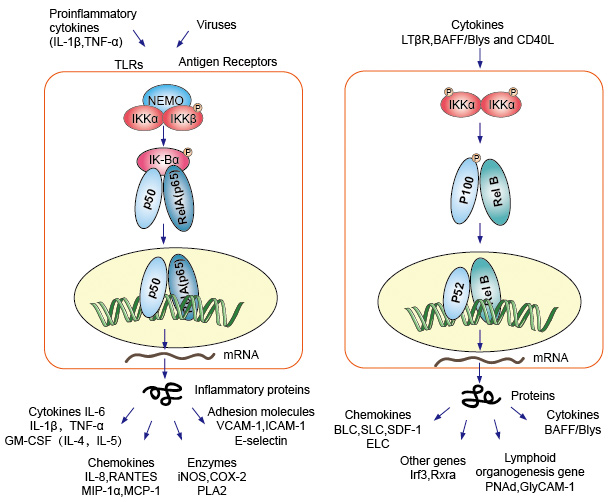
Figur 4. Down-stream signalering av NF-kB signaliserer veien.
5. Forholdet med sykdommer.
- inflammatorisk grunnlag av metabolske sykdommer.,
Det IKK/NF-kB signaliserer vei er nøkkelen til kobling av metabolisme, betennelse, og insulin action. De fleste av metabolsk stress signaler som resulterer i insulin resistens eller bukspyttkjertelen β celle dysfunksjon, enten forårsaket av intracellulære eller ekstracellulære signaler, møtes på NF-kB-aktivere kinase IKKb og andre store inflammatorisk kinase, JNK-mitogen-aktivert protein kinase (MAPK).
- NF-kB og glykolysen.
Glykolysen kan generere ATP i større mengder, og i et raskere tempo enn oksidativt phosphorylation., Glukose er derfor et viktig næringsstoff for kreft og normal voksende celler. NF-kB-koordinater mange av de signaler som stasjonen celle aktivering og spredning under immunitet, betennelse, og onkogenese.
- NF-kB og oksidativ metabolisme.
NF-kB styrer energi homeostase og metabolske tilpasninger ved å kontrollere balansen mellom glykolysen og åndedrett for energiforsyning. Hemming av NF-kB/RelA av ulike virkemidler redusert oksygen forbruk og forårsaket omprogrammering aerobic glykolysen