Figura 1. Calea de semnalizare canonică și non-canonică NF-kB
ce este NF-kB? NF-kB este un nume scurt al factorului Nuclear kappa-light-chain-enhancer al celulelor B activate. Nu este o singură proteină, ci o mică familie de factori de transcripție inductibili care joacă un rol important în aproape toate celulele mamiferelor. Controlează transcripția ADN, producția de citokine, supraviețuirea celulară și alte evenimente celulare importante, în special joacă un rol-cheie în reglarea răspunsului imun la infecție.NF-kB molecular sunt de obicei dimeri., O structură tipică a NF-kB este dimerul P50-P65 (NF-KB1/RelA). Formarea dimerului este necesară pentru legarea ADN-ului, doi monomeri NF-kB se leagă de ADN ca dimer. Regiunile n-terminale ale dimerului sunt responsabile pentru contactul ADN specific. Regiunile C-terminale sunt de obicei foarte conservate, ele sunt responsabile pentru dimerizarea și contactul nespecific cu fosfatul ADN. Întreaga moleculă NF-kB este la fel ca o menghină de clește pe lanțul ADN și funcționează ca factor de transcripție (figura 2).
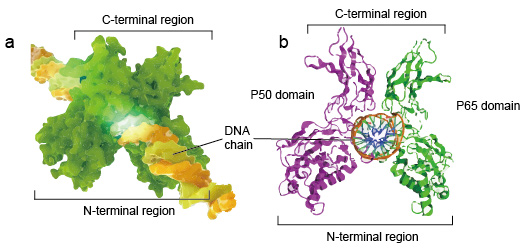
Figura 2. Structura legării dimerului proteic NF-kB cu lanțul ADN.,
familia NF-kB
deoarece membrii familiei NF-kB împărtășesc omologia structurală cu oncoproteina retrovirală v-Rel. așa că au numit și proteine NF-kB / Rel. NF-kB familia factorului de transcriere la mamifere este format din 5 proteine: p65/RelA, RelB, c-Rel, p105/p50 (NF-kB1), și p100/p52 (NF-kB2). Aceste 5 proteine se asociază între ele și formează complexe homo/heterodimerice active transcriptional distincte (Figura 3). Toate au un domeniu comun conservat rel omologie (RHD) de 300 aa lungime., Aceste domenii RHD au funcții multiple, cum ar fi dimerizarea, legarea ADN-ului, interacțiunea cu IkBs și translocația nucleară. Deși membrii familiei de proteine NF-kB pot forma până la 15 dimeri diferiți, dar mulți dintre ei nu au demonstrat încă. Cea mai abundentă formă de dimer NF-kB este heterodimerul p50/p65 care a fost fond în aproape toate tipurile de celule. Deoarece numai P65 / Rel, RelB și C-Rel au domenii de transactivare carboxi-terminale( TAD), familia de proteine NF-kB poate fi împărțită în continuare în două grupe., p50 și p52 sunt generate prin prelucrarea moleculelor precursoare p105 și, respectiv, P100. Prin urmare, nu toate combinațiile de dimeri Rel sunt active transcriptional.
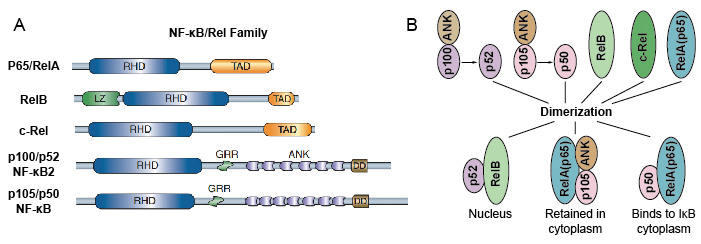
Figura 3. Proteina 5 și complexele lor homo / heterodimerice din familia NF-kB.NF-kB variatoare de proteine ca factor de transcriere nucleară, ei trebuie să migreze la nucleu, combinat cu ADN-ul pentru a avea funcția. În cele mai multe tipuri de celule normale în stare de repaus, NF-KB a fost inactiv și reține în citoplasmă., Acestea se leagă de un inhibitori specifici numiți proteină IK-B, care s-ar putea lega de domeniul omologiei Rel (RHD) al NF-kB și interferează cu funcția sa de secvență de localizare nucleară (NLS). Aceste inhibitor de proteine, care includ IkBa, IkBb și IkBg, conține 6-7 ankyrin repetă care mediază legarea la RHD. Aceste repetări sunt, de asemenea, prezente în C-terminal jumătăți ale NF-kB2/p100 și NF-kB1/p105 precursori, care, de asemenea, funcționează ca IkBs și păstreze partenerii lor, Rel proteine, în citoplasmă., Pentru a activa molecula NF-kB, celulele trebuie mai întâi să separe proteina NF-kB de inhibitorii lor. Există două căi majore de semnalizare care conduc la disocierea inhibitorului proteinei IK-B de la dimmerul NF-kB și permit translocarea dimerilor NF-kB din citoplasmă în nucleu.
1., Canonic/Clasică Cascadă
Canonice sau clasice cascadă de semnalizare incepand de la suprafața celulei receptorilor de citokine pro-inflamatorii și pathogen-associated molecular patterns (PAMPs), cum ar fi factorul de necroză tumorală receptorilor (TNFR), toll-like receptorilor (TLR) și T/B cell receptor. Acești receptori care se leagă cu moleculele lor de ligand și transferă semnalul peste membrana celulară, determină activarea complexului IkB kinase (IKK). Cea mai comună formă de acest complex format heterodimer de IKKa și IKKß subunități catalitice și o IKKy de reglementare subunitate., Unitatea IKKy, de asemenea, numit NEMO pentru NF-kB modulator esențial. Activat IKK complex, predominant acționează prin IKKß într-un IKKy manieră dependentă, catalizează fosforilarea IkBs (la site-uri echivalente cu Ser32 și Ser36 de IkBa), polyubiquitination (la site-uri echivalente cu Lys21 și Lys22 de IkBa) și degradarea ulterioară de 26S proteazomal. Dimerii NF-kB eliberați (cel mai frecvent dimerul p50–RelA), se translocează în nucleu, leagă ADN-ul și activează transcripția genei în jos.
2., Calea non-canonică/alternativă cascadă
o altă cale de activare NF-kB numită cale no-canonică sau alternativă. Această cale este independentă de IKKß și IKKy, dar depinde de IKKa dimmer în schimb. Transferul de semnalizare în citoplasmă prin receptorul LT-β sau BAFF. Fosfonați o proteină NIK, proteina NIK apoi fosfonați Homodimerii IKKa. Obiectivul pentru Homodimerii IKKa în această cale este NF-kB2 / p100, care este fosforilat în două locuri terminale C., Fosforilarea acestor situri este esențială pentru prelucrarea p100 la p52, care este, de asemenea, dependentă de poliubichitinare și degradare proteazomală. Cu toate acestea, în loc de a conduce pentru a finaliza p100 degradare, cum se vede cu IkBs, fosforilarea dependente de ubiquitination din p100 rezultate doar în degradarea său inhibitor C-terminal jumătate părți. Odată ce jumătatea terminală C este degradată, porțiunea N-terminală a NF-kB (polipeptida p52 care conține RHD) este eliberată., Ca RHD p100 este cel mai frecvent asociat cu RelB, activarea acestei „alternative” calea rezultate în translocarea nucleară de p52 RelB dimeri. Dimerul se leagă în cele din urmă de ADN și activează transcripția genei în jos.
3. Ubiquitina este o proteină 76-amino-acidă care este foarte conservată și exprimată omniprezent în toate eucariotele de la drojdie la om. Acidul carboxilic din glicina C-terminală a ubiquitinei poate fi atașat covalent la Amina epsilon a lizinei pe o altă proteină printr-o legătură izopeptidică. Acest proces se numește „ubiquitination”., Mai mult decât atât, C-terminal coada de ubiquitin poate fi atașat direct la terminal N metionină de un alt ubiquitin pentru a forma liniară polyubiquitin lanț. Ubicuitatea joacă un rol esențial în reglarea căilor NF-kB. În celulele NF-kB nestimulate, se leagă de proteinele inhibitoare ale familiei kB (IkB) și este sechestrat în citoplasmă., La stimulare, IkB este fosforilată de IkB-kinazei (IKK) complexe, Fosforilată IkB este ulterior ubiquitinated și degradate de 26 de proteazom, permițând, astfel, NF-kB, pentru a transloca la nucleu, unde se regleaza expresia de o serie de gene. Ca subunitate de reglementare a complexului IKK, NEMO a fost propus a fi factorul cheie pentru transducerea semnalului de ubicuitate la activarea IKK. Mai multe DUBs funcționează ca regulatoare cheie negative ale IKK pentru a permite un control strict al activării NF-kB., Una dintre cele mai bune studiat DUBs este A20, S-a propus ca A20 suprima hyperactivation de NFkB de deubiquitinating K63-legate polyubiquitin atașat la RIP1 prin N-terminal OTU domeniu, precum și promovarea K48-legate polyubiquitination de RIP1 pentru proteasomal degradare.
4. NF-kB acționează ca un mediator central al răspunsurilor imune și inflamatorii și este implicat în răspunsurile la stres și reglarea proliferării celulare și a apoptozei. Genele țintă NF-kB respective permit organismului să răspundă eficient la aceste schimbări de mediu., Calea clasică NF-kB este activată de o varietate de semnale inflamatorii, rezultând expresia coordonată a mai multor gene imune inflamatorii și înnăscute. Citokinele proinflamatorii IL-1b și TNF-a activează NF-kB, iar expresia lor este indusă ca răspuns la activarea NF-kB, formând astfel o buclă de amplificare înainte. Calea alternativă pentru NF-kB rezultate în translocarea nucleară de p52 RelB dimeri, este strict dependentă de IKKa homodimers și este activat de LTbR, BAFF și CD40L de NIK., Multe date sugerează cu tărie că calea alternativă joacă un rol central în exprimarea genelor implicate în dezvoltarea și întreținerea organelor limfoide secundare. Factorii de transcripție ai familiei NF-kB controlează expresia unui număr mare de gene țintă ca răspuns la schimbările din mediu, contribuind astfel la orchestrarea răspunsurilor inflamatorii și imune.
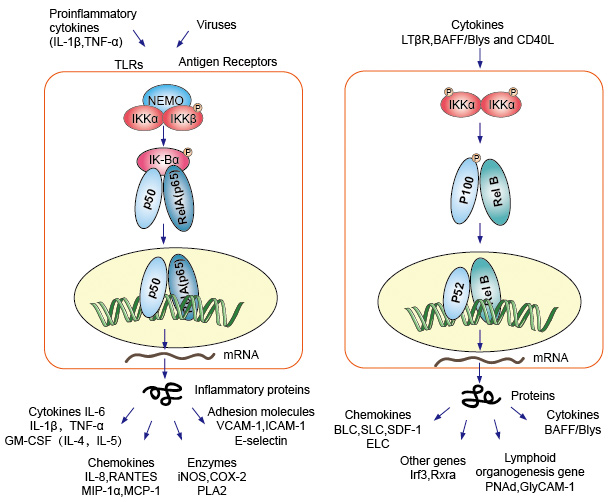
Figura 4. Semnalizarea în flux a căii de semnalizare NF-kB.
5. Relația cu bolile.
- baza inflamatorie a bolilor metabolice.,calea de semnalizare IKK/NF-kB este esențială pentru legarea metabolismului, inflamației și acțiunii insulinei. Cele mai multe dintre stres metabolic semnalele care rezultă în rezistența la insulină sau pancreatice β celulare disfuncție, fie induse de intracelular sau extracelular indicii, converg pe NF-kB-activarea kinazei IKKb și alte mari inflamatorii kinazei, EDUARD-mitogen activated protein-kinaza (MAPK).
- NF-kB și glicoliză.glicoliza poate genera ATP în cantități mai mari și într-un ritm mai rapid decât fosforilarea oxidativă., Prin urmare, glucoza este un nutrient esențial pentru cancer și celulele proliferante normale. NF-kB coordonează multe dintre semnalele care conduc activarea și proliferarea celulelor în timpul imunității, inflamației și oncogenezei.
- NF-kB și metabolismul oxidativ.NF-kB guvernează homeostazia energetică și adaptarea metabolică prin controlul echilibrului dintre glicoliză și respirație pentru furnizarea de energie. Inhibarea NF-kB/RelA prin diferite mijloace a diminuat consumul de oxigen și a determinat reprogramarea glicolizei aerobe
- NF-kB și glicoliză.glicoliza poate genera ATP în cantități mai mari și într-un ritm mai rapid decât fosforilarea oxidativă., Prin urmare, glucoza este un nutrient esențial pentru cancer și celulele proliferante normale. NF-kB coordonează multe dintre semnalele care conduc activarea și proliferarea celulelor în timpul imunității, inflamației și oncogenezei.