kuva 1. Kanoninen ja ei-kanoninen NF-kB signalointireitti
mikä on NF-kB?
NF-kB on aktivoitujen B-solujen ydintekijän kappa-valoketjun tehostaja. Se ei ole yksittäinen proteiini, mutta pienen perheen indusoituva transkriptio tekijät että pelata tärkeä rooli lähes kaikissa nisäkässoluissa. Se ohjaus DNA transkriptio, sytokiinien tuotantoa, solujen eloonjäämistä ja muita tärkeitä solun tapahtumia, erityisesti, on keskeinen rooli säätelyssä immuunivastetta infektio.
NF-kB molekyylit ovat yleensä dimeerejä., NF-kB: n tyypillinen rakenne on P50-P65 dimeeri (NF-kB1/RelA). Dimeerimuodostus on välttämätön DNA: n sitoutumiselle, kaksi NF-kB-monomeeria sitoutuu DNA: han dimeerinä. Dimerin n-terminaalialueet vastaavat spesifisestä DNA-kontaktista. C-terminaali alueet ovat yleensä hyvin säilynyt, he ovat vastuussa dimerization ja epäspesifinen DNA-fosfaatti yhteyttä. Koko NF-kB-molekyyli on aivan kuin Pihtipudas DNA-ketjussa ja toimii transkriptiokertoimena (kuva 2).
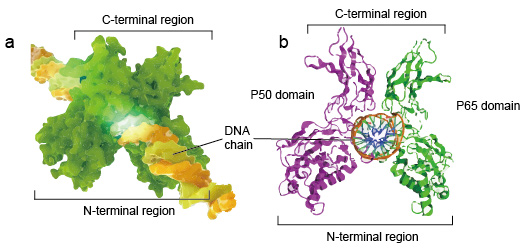
Kuva 2. NF-kB-proteiinidimeerin rakenne, joka sitoutuu DNA-ketjuun.,
NF-kB-perhe –
Koska NF-kB perheen jäsenet jakavat rakenteellinen homologia kanssa retroviral oncoprotein v-Rel. niinpä he kutsuivat myös NF-kB/Rel-proteiineja. NF-kB transkriptio tekijä perheen nisäkkäillä koostuu 5 proteiinit: p65/Suh, RelB, c-Rel, p105/p50 (NF-kB1), ja p100/p52 (NF-ks2). Nämä 5 proteiinit yhdistää toisiinsa ja muodostavat erillisiä transcriptionally aktiivinen homo/heterodimeric komplekseja (Kuva 3). Niillä kaikilla on yhteinen säilytetty Rel homology domain (RHD), jonka pituus on 300 aa., Nämä RHD domain on useita toimintoja, kuten dimerization, DNA: ta sitova, vuorovaikutus IkBs, ja ydinvoima translokaatio. Vaikka NF-kB proteiini perheen jäsenet voivat muodostaa jopa 15 eri dimeerit, mutta monet heistä ei ole vielä osoitettu. NF-kB dimerin runsain muoto on P50 / p65 heterodimeeri, jota on rahastoitu lähes kaikissa solutyypeissä. Koska vain P65/Rel, RelB ja c-Rel on karboksi-terminaali transactivation verkkotunnukset (TAD), NF-kB-perheen proteiinit voidaan jakaa edelleen kahteen ryhmään., P50 ja p52 syntyvät prekursorimolekyylien P105 ja p100 prosessoinnista. Siksi kaikki Rel-dimeerien yhdistelmät eivät ole transkriptioaktiivisia.
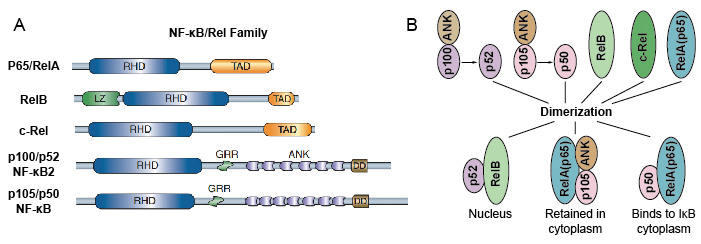
Kuva 3. NF-kB-suvun 5 proteiinia ja niiden homo / heterodimeerisiä komplekseja.
NF-kB signalointi koulutusjakso,
NF-kB proteiini himmentimet kuin ydinvoima transkriptio tekijä, ne täytyy siirtyä ydin, yhdistettynä DNA-on-toiminto. Useimmissa normaaleissa solutyypeissä lepotilassa NF-KB oli inaktiivinen ja säilyi sytoplasmassa., Ne sitovat tietyn estäjiä kutsutaan IK-B-proteiini, joka voi sitoutua Rel homology domain (RHD) NF-kB ja häiritä sen ydinvoiman lokalisointi järjestyksessä (NLS) – toiminto. Nämä estäjä proteiineja, jotka sisältävät IkBa, IkBb ja IkBg, sisältää 6-7 ankyrin toistaa, että välittäjänä sitova RHD. Nämä toistot ovat läsnä myös C-päätteen puoliskot NF-ks2/p100 ja NF-kB1/p105 esiasteita, jotka toimivat myös IkBs ja säilyttää heidän kumppaneilleen, Rel proteiineja, sytoplasmassa., NF-kB-molekyylin aktivoimiseksi solujen on ensin erotettava NF-kB-proteiini inhibiittoreistaan. On olemassa kaksi suurta signaalinvälitysreittien johtaa IK-B-proteiini estäjä irtaantuu NF-kB himmennin ja anna translokaatio NF-kB-dimeerit siitä tulee ydin solulimaan.
1., Kanoninen/Klassinen Cascade
Kanoninen tai klassisen signalointi cascade alkaen solun pinnan reseptorin pro-inflammatoristen sytokiinien ja pathogen-associated molecular patterns (PAMPs), kuten tuumorinekroositekijä reseptorin (TNFR), toll-kaltainen reseptori (TLR) ja T/B-solujen reseptori. Nämä reseptorit sitovat niiden ligandi molekyylejä ja siirtää signaalin solukalvon läpi, koska aktivointi IkB-kinaasin (IKK) monimutkainen. Tämän kompleksin yleisin muoto on IKKa-ja IKKß-katalyyttisten alayksiköiden heterodimeeri ja IKKy regulatory-alayksikkö., IKKy-yksikkö kutsui Nemoa myös NF-kB essential modulaattoriksi. Aktivoitu IKK monimutkainen, pääasiassa kautta toimivat IKKß on IKKy-riippuvaisella tavalla, katalysoi fosforylaatio ja IkBs (at sivustoja vastaa Ser32 ja Ser36 ja IkBa), polyubiquitination (at sivustoja vastaa Lys21 ja Lys22 ja IkBa) ja myöhempi hajoaminen, jonka 26S-proteasomin. Julkaistu NF-kB-dimeerit (yleisimmin p50–Riippuu dimeeri), translocate tumaan, sitovat DNA: ta ja aktivoi alas-stream-geenin transkriptio.
2., Ei-kanoninen/Vaihtoehtoinen polku Cascade
Toinen NF-kB aktivoimalla koulutusjakson kutsutaan ei-kanoninen tai vaihtoehtoinen reitti. Tämä polku on itsenäinen Ikkßistä ja Ikkystä, mutta sen sijaan riippuvainen IKKa himmeristä. Signalointi siirtyy sytoplasmaan LT-β-tai BAFF-reseptorin kautta. Fosfonaatti NIK-proteiini, NIK-proteiini ja sen jälkeen Fosfonaatti IKKa-homodimmerit. IKKa-homodimeerien kohde tällä reitillä on NF-kB2/p100, joka fosforyloidaan kahdessa C-terminaalissa., Fosforylaation näistä sivustoja on välttämätöntä p100 käsittely p52, joka on myös riippuvainen polyubiquitination ja proteasomal hajoamista. Kuitenkin, sen sijaan, mikä johtaa täydellinen p100 hajoamista, kuten IkBs, että fosforylaation-riippuvainen ubiquitination ja p100 johtaa vain heikkeneminen sen estävä C-terminaali puoli osia. Kun C-terminaalinen puolikas hajoaa, vapautuu NF-kB: n n-terminaalinen osa (RHD: tä sisältävä P52-polypeptidi)., Koska RHD ja p100 on yleisimmin liitetty RelB, aktivointi tämä ”vaihtoehtoinen” polku johtaa ydinaseiden translokaatiossa p52–RelB dimeerit. Dimeeri sitoutuu lopulta DNA: han ja aktivoi down stream-geenin transkription.
3. Reitin asetus
Ubikitiinipromoottori on 76-aminohappo proteiini, joka on hyvin säilynyt ja kaikkialle ilmaistu kaikki eukaryooteissa hiivasta ihmiseen. Karboksyylihapoksi C-terminaali glysiini ubikitiinipromoottori voidaan kovalenttisesti kiinnitetty epsilon amiini lysiini-on toinen proteiinin kautta isopeptide bond. Tätä prosessia kutsutaan ”ubikitinaatioksi”., Lisäksi, C-terminaali häntä ubikitiinipromoottori voi olla suoraan kiinni N-terminaali metioniini toisen ubikitiinipromoottori muodostaa lineaarinen polyubiquitin ketju. Ubikitinaatiolla on keskeinen rooli NF-kB-reittien säätelyssä. Vuonna stimuloimattomissa soluissa, NF-kB sitoutuu estävä proteiineja kB perhe (IkB) ja on eristäytynyt sytoplasmassa., Kun stimulaatio, IkB on fosforyloitu, jonka IkB-kinaasin (IKK) monimutkainen, Fosforyloitu IkB on myöhemmin ubiquitinated ja hajoavan 26S-proteasomin, jolloin NF-kB translocate tumaan, jossa se säätelee ilmaus lukuisia geenejä. Koska sääntely-alayksikköön IKK monimutkainen, NEMO on ehdotettu olevan keskeinen tekijä transducing ubiquitination signaalin aktivointi IKK. Useita DUBs toimivat avain negatiivinen sääntelyviranomaisten IKK jotta tiukka valvonta NF-kB aktivaatiota., Yksi parhaista tutkittu DUBs on A20, Se on ehdotettu, että A20 tukahduttaa hyperactivation ja Ne, joita deubiquitinating K63-liittyvät polyubiquitin kiinnitetty RIP1 kautta N-terminaali OTU domain, sekä edistää K48-liittyvät polyubiquitination ja RIP1 varten proteasomal hajoamista.
4. Loppupään signalointi
NF-kB toimii keskeisenä välittäjänä immuuni-ja tulehduksellinen vastauksia, ja on mukana stressistä ja sääntelyn solujen proliferaation ja apoptoosin. Vastaavat NF-kB-kohdegeenit mahdollistavat organismin tehokkaan reagoinnin näihin ympäristömuutoksiin., Klassisen NF-kB reitti on käytössä erilaisia tulehduksellinen signaaleja, jolloin koordinoida ilmaus useita tulehduksellinen ja synnynnäisen immuniteetin geenejä. Myös proinflammatoristen sytokiinien IL-1b ja TNF-a aktivoi NF-kB, ja niiden ilme on aiheuttama vastauksena NF-kB aktivaatiota, muodostaen vahvistavaa rehun eteenpäin silmukka. Vaihtoehto koulutusjakson NF-kB tulokset ydinvoima-translokaation p52–RelB dimeerit, on tiukasti riippuvainen IKKa homodimers ja aktivoidaan LTbR, BAFF ja CD40L by NIK., Monet tiedot viittaavat vahvasti siihen, että vaihtoehtoinen reitti on keskeinen rooli geenien mukana kehittäminen ja ylläpito keskiasteen lymfaattisessa elimet. Transkriptio tekijät NF-kB perheen ohjaus ilmaus suuri määrä kohde-geenien vastauksena ympäristön muutoksiin, mikä auttaa ohjaamaan inflammatorisia ja immuunivastetta.
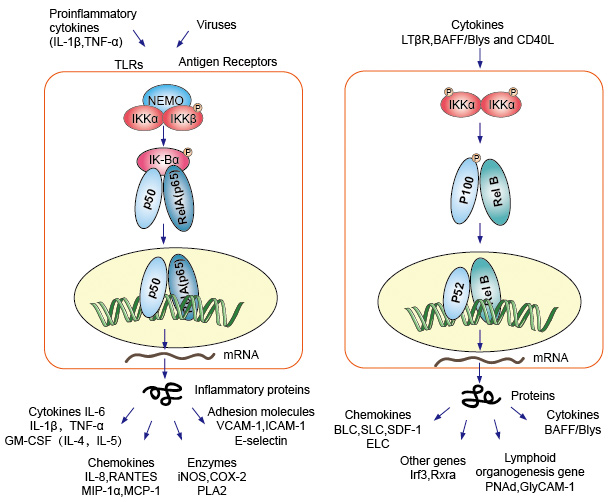
Kuva 4. NF-kB-signalointireitin alasvirtaus.
5. Suhde sairauksiin.
- metabolisten sairauksien tulehduspohja.,
IK / NF-kB-signalointireitti on avain aineenvaihdunnan, tulehduksen ja insuliinin vaikutuksen yhdistämiseen. Suurin osa metabolista stressiä signaaleja, jolloin insuliiniresistenssi tai haiman β-solujen toimintahäiriö, onko aiheuttama solunsisäisen tai solunulkoisen vihjeet, lähentymässä NF-kB-aktivointi kinaasi IKKb ja muut suuret tulehduksellinen kinaasi, JNK-mitogeeni aktivoitua proteiinikinaasia (MAPK).
- NF-kB ja glykolyysi.
Glykolyysin voi tuottaa ATP suurempia määriä ja nopeammin kuin oksidatiivisen fosforylaation., Siksi glukoosi on tärkeä ravintoaine syövälle ja normaaleille proliferoiville soluille. NF-kB koordinoi monia signaaleja, jotka ajavat solujen aktivoitumista ja proliferaatiota immuniteetin, tulehduksen ja onkogeneesin aikana.
- NF-kB ja oksidatiivinen aineenvaihdunta.
NF-kB säätelee energian homeostaasia ja metabolista sopeutumista säätelemällä glykolyysin ja hengityksen välistä tasapainoa energiansaannin kannalta. NF-kB/Relan estyminen eri tavoin vähensi hapenkulutusta ja aiheutti aerobisen glykolyysin uudelleenohjelmoinnin